Соматосенсорные вызванные потенциалы (ССВП) головного мозга
В неврологии есть такой диагностический метод, как регистрация вызванных потенциалов. Соматосенсорные ВП – это обследование, которое позволяет врачу оценить ответ от центральной нервной системы (в которую входят спиной и головной мозг) на стимуляцию волокон периферической части нервной системы. По тому, когда и как проявляется этот ответ, можно определить наличие неврологической патологии, оценить ее характер и тяжесть. С развитием технологий это исследование стало активно применяться в диагностике самых разных заболеваний неврологического профиля.
20 — 50 минут
(продолжительность процедуры)
Амбулаторно
Диагностика нарушений с помощью ССВП
Существует два вида стимуляции: верхних конечностей и нижних. Методики различаются местами наложения электродов. При наличии показаний можно провести стимуляцию практически любого нерва, относящегося к периферической нервной системе. Количество и положение электродов зависят от целей исследования. При подаче сигналов врач с помощью оборудования оценивает целый ряд показателей, которые затем сравнивает с нормативными.
При подаче сигналов врач с помощью оборудования оценивает целый ряд показателей, которые затем сравнивает с нормативными.
Стимуляция верхних конечностей происходит чаще всего с помощью наложения электрода на запястье.
При стимуляции нижних конечностей суть исследования заключается в воздействии на нервы, соответственно, ног. Электрод, подающий импульсы, обычно располагают в области лодыжки.
ССВП позволяет оценить наличие нарушение в периферической или центральной нервной системе. В некоторых случаях метод позволяет довольно четко определить место поражения. Метод специфический, поэтому обследование проводится совместно с другими мероприятиями по комплексной диагностике.
Показания
- Оценки уровня демиелинизации. Позволяет подтвердить повреждение спинного мозга. Не применяется для дифференциальной диагностики, так как есть заболевания, которые дают похожие результаты.
- Диагностика функциональной потери чувствительности – метод подтверждает соматическую сохранность нервных проводящих путей, что позволяет поставить диагноз функциональной недостаточности.

- Исследование проводят в рамках оценки состояния спинного мозга при травмах и нарушении кровообращения спинного мозга.
- Определение степени поражения спинного мозга при миелопатии (например, на фоне грыжи межпозвонкового диска).

Как проходит обследование
Сначала врач накладывает электроды в соответствии с выбранной схемой исследования, их может быть достаточно много. Типичные места для прикрепления стимулирующих элементов – это лодыжки, запястья и колени. Электроды, благодаря которым происходит регистрация отклика, закрепляют на голове и спине, в зависимости от того, с какой зоны головного мозга необходимо получить информацию.
Непосредственно обследование начинается с подачи стимулирующих сигналов. Под воздействием импульсов периферические нервы вызывают сокращение мышц в соответствующей зоне. Минимальное количество стимулов для успешного исследования – не менее 200–500, в зависимости от целей и задач обследования.
С помощью компьютера происходит расчет среднего времени, за которое сигнал достигает головной мозга.
Соматосенсорные вызванные потенциалы – это высокочувствительное исследование, на которое может повлиять множество факторов. Поэтому, если результат оказывается недостаточно информативным, обследование повторяют еще раз.
Врач, выполняющий ССВП
Часто задаваемые вопросы
Какие заболевания можно выявить методом регистрации ССВП?
- рассеянный склероз
- энцефаломиелит, миелит и их последствия
- плексопатии, полинейропатии
- нарушения потенции и мочеиспускания
- хронические болевые синдромы
- травмы и сосудистые заболевания спинного мозга, головного мозга и их последствия
Регистрация ССВП часто дополняется проведением игольчатой и стимуляционной электронейромиографии (ЭНМГ).
Исследование не используется для дифференциальной диагностики, применяется для оценки функционального состояния проводящих путей. Таким образом, окончательная постановка диагноза не производится только по результатам ССВП. Метод применяется в комплексной диагностике.
Метод применяется в комплексной диагностике.
Насколько болезненна процедура регистрации ССВП?
Процедура совершенно безболезненная. Некоторый дискомфорт могут представлять ощущения от подергивания мышц. Сильная стимуляция не требуется, поэтому боли не возникает.
Как подготовиться к процедуре?
Необходимо вымыть голову, желательно не проводить укладку волос или выполнять сложную прическу – электроды будут накладывать и на волосистую часть головы.
Есть ли противопоказания?
Противопоказаний нет.
Сколько длится процедура?
От 20 до 50 минут, в зависимости от количества импульсов. Обработка результатов может занять некоторое время.
Где можно пройти обследование?
В клинике «ЦЭЛТ» профессиональные диагносты работают на современном оборудовании – а это значит, что результаты будут максимально точными. Можно записаться на удобное время, получить предварительную консультацию от профильного специалиста. Быть здоровым легко и просто со специалистами многофункциональной клиники «ЦЭЛТ».
На вопросы отвечает:
Вызванные потенциалы мозга
Вызванные потенциалы мозга (ВП) – тестирование функции зрительного, акустического и соматосенсорного анализаторов. Исследуются когнитивные функции на основе эндогенного когнитивного вызванного потенциала, и исследуется центральное и периферическое звено вегетативной нервной системы с помощью вызванного кожного симпатического потенциала.
Производится оценка высших корковых функций мозга человека, таких как распознавание стимула, дифференцировка, запоминание и мыслительные процессы, связанные с принятием решения.
|
|
Показания к обследованию:
|

Соматосенсорные вызванные потенциалы.
В ходе этого исследования проводится:
- оценка сохранности афферентных путей при диагностике поражений спинного и головного мозга, периферических нервов;
- определение тяжести повреждения мозга и возможных последствий травмы и комы;
|
Показания к обследованию:
|
Акустические стволовые вызванные потенциалы.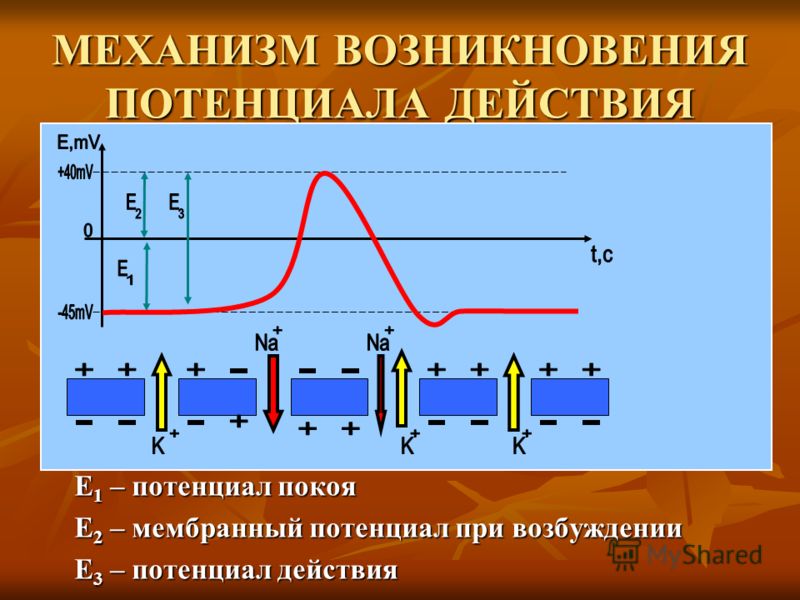
Проводится оценка функционального состояния периферических и стволовых акустических структур.
Показания к обследованию:
|
Зрительные вызванные потенциалы.
Оценка состояния зрительного нерва, хиазмы, зрительного тракта и зрительной коры, остроты зрения и ее корригируемости, зрительных нарушений и их динамики при лечении.
Показания к обследованию:
|

Вызванные кожные симпатические потенциалы – оценка функции вегетативной нервной системы.
Показания к применению:
- дифференциальная диагностика нарушений вегетативной регуляции у больных неврологического и терапевтического профиля и вегетативных полинейропатий;
- оценка влияния неблагоприятных факторов профессиональной деятельности, индивидуальный подбор фармакотерапии вегетативных кризов, нейроциркуляторной дистонии.
Потенциал. Разность потенциалов. Напряжение.Эквипотенциальные поверхности
Основные ссылки
CSS adjustments for Marinelli theme
Объединение учителей Санкт-Петербурга
Форма поиска
Поиск
Вы здесь
Главная » Потенциал. Разность потенциалов. Напряжение…
| Потенциал. Разность потенциалов. | |
Потенциал электростатического поля — скалярная величина, равная отношению потенциальной энергии заряда в поле к этому заряду: — энергетическая характеристика поля в данной точке. Потенциал не зависит от величины заряда, помещенного в это поле. | |
Т.к. потенциальная энергия зависит от выбора системы координат, то и потенциал определяется с точностью до постоянной. За точку отсчета потенциала выбирают в зависимости от задачи: а) потенциал Земли, б) потенциал бесконечно удаленной точки поля, в) потенциал отрицательной пластины конденсатора. |
|
— следствие принципа суперпозиции полей (потенциалы складываютсяалгебраически | |
Потенциал численно равен работе поля по перемещению единичного положительного заряда из данной точки электрического поля в бесконечность. В СИ потенциал измеряется в вольтах: |
|
Разность потенциалов | |
| |
Напряжение — разность значений потенциала в начальной и конечнойточках траектории. Напряжение численно равно работе электростатического поля при перемещении единичного положительного заряда вдоль силовых линий этого поля. Разность потенциалов (напряжение) не зависит от выбора системы координат! | |
Единица разности потенциалов
Напряжение равно 1 В, если при перемещении положительного заряда в 1 Кл вдоль силовых линий поле совершает работу в 1 Дж. | |
Связь между напряженностью и напряжением. | |
Из доказанного выше: → напряженность равна градиенту потенциала (скорости изменения потенциала вдоль направления d). | |
Из этого соотношения видно:
| |
Эквипотенциальные поверхности. ЭПП — поверхности равного потенциала. Свойства ЭПП: — работа при перемещении заряда вдоль эквипотенциальной поверхности не совершается; — вектор напряженности перпендикулярен к ЭПП в каждой ее точке. | |
| |
Измерение электрического напряжения (разности потенциалов) Между стержнем и корпусом — электрическое поле. Измерение потенциала кондуктора Измерение напряжения на гальваническом элементе Электрометр дает большую точность, чем вольтметр. | |
Потенциальная энергия взаимодействия зарядов. |
|
Потенциал поля точечного заряда |
|
| |
Потенциал заряженного шара а) Внутри шара Е=0, следовательно, потенциалы во всех точках внутри заряженного металлического шара одинаковы (!!!) и равны потенциалу на поверхности шара. б) Снаружи поле шара убывает обратно пропорционально расстоянию от центра шара, как и в случае точечного заряда. | |
Перераспределение зарядов при контакте заряженных проводников. Переход зарядов происходит до тех пор, пока потенциалы контактирующих тел не станут равными. |
|
 Напряжение.
Напряжение.


Теги:
конспект
Потенциал развития есть у каждого региона России
Завтра, 12 июня День России. День, который пока еще, считаю, не занял в сознании подавляющего числа россиян места, соответствующего его историческому значению. Уверена, что год за годом мы все полнее будем раскрывать его потенциал, отдавая должное сделанному выбору и общему импульсу, полученному в тот переломный момент.
Потому что эта дата одновременно и важнейшая точка отсчёта, и символ начала глубинных преобразований, вернувших
наше Отечество в экономическое и социальное пространство, в котором
существовало, продолжает существовать и развиваться подавляющее большинство
человечества. Конечно, как всегда в переломные моменты, нам пришлось нелегко. Порой
просто трудно, тяжело. Тем более, что нашего возвращения как равных партнеров
ждали далеко не все. И с самого начала на этом пути нам пришлось столкнуться с проявлениями недоброжелательности, противодействия на международной арене. Со временем такое сопротивление не только не ослабло, а, напротив, как мы видим,
для группы западных стран сложилось в систему, в цельную и уже не скрываемую
стратегию сдерживания России.
Конечно, как всегда в переломные моменты, нам пришлось нелегко. Порой
просто трудно, тяжело. Тем более, что нашего возвращения как равных партнеров
ждали далеко не все. И с самого начала на этом пути нам пришлось столкнуться с проявлениями недоброжелательности, противодействия на международной арене. Со временем такое сопротивление не только не ослабло, а, напротив, как мы видим,
для группы западных стран сложилось в систему, в цельную и уже не скрываемую
стратегию сдерживания России.
Но несмотря ни на что, за эти годы мы сумели четко определить национальные интересы, заявили о них и твердо отстаиваем свой суверенитет и свое право. Мы стали сильнее. Россия восстановилась в роли державы, без учёта позиции которой невозможно решить ни один хоть сколько‑нибудь важный вопрос международных отношений, мировой политики и экономики.
Сегодня Российская
Федерация — страна, у которой есть идея, консолидирующая общество, объединяющая
народы в единую нацию. Эта идея – патриотизм, любовь к Отечеству, историческая
и гражданская идентичность россиян, в основе которой проверенные временем
общественно-политические, моральные, духовные ценности. Мы умеем дорожить своей
великой историей, сохранять связь между поколениями, лучшие традиции и богатейшее наследие нашего народа.
Мы умеем дорожить своей
великой историей, сохранять связь между поколениями, лучшие традиции и богатейшее наследие нашего народа.
У нас в хорошем смысле амбициозные планы – осуществить в исторически короткие сроки прорыв на самые высокие рубежи технологического, экономического, социального, культурного развития, обеспечить достойное качество жизни всем российским гражданам. Планы реальные, главные показатели которых запечатлены в Национальных целях развития на период до 2030 года. На их достижение направлены национальные проекты, государственные программы, федеральные и региональные стратегии развития.
Считаю важным в День России еще раз обратить внимание на то, без учета чего невозможна
реализация желаемого для нас сценария. Это, конечно, региональный аспект.
Региональный потенциал. Он способен стать нашим абсолютно уникальным
преимуществом, если мы сумеем сохранить, объединить и использовать ресурс всего
нашего природного, пространственного и национального многообразия! Для России —
федеративного государства — как ни для какой другой страны в мире в осуществлении любых планов велика роль регионов. Каждого из 85 субъектов. Этот
акцент по‑новому обозначил недавно в своем выступлении на ПМЭФ Владимир
Владимирович Путин.
Каждого из 85 субъектов. Этот
акцент по‑новому обозначил недавно в своем выступлении на ПМЭФ Владимир
Владимирович Путин.
Президент поставил конкретные задачи по созданию условий для развития бизнеса в регионах, планирования экономической деятельности на основе понятной информации, в предсказуемой, дружественной для бизнеса обстановке. Каждый субъект Федерации должен четко и открыто представить приоритетные направления своего развития, в том числе градостроительные, инфраструктурные планы. Устранить избыточные формальности и согласования. Разработать понятный алгоритм действий, чтобы сократить время от бизнес-идеи до открытия нового производства.
Понятно, насколько
важно в этой работе обеспечить слаженность совместных действий Правительства и глав, органов государственной власти регионов. Естественно, в этой сфере
широкое поле деятельности и для нас, парламентариев. Особенно для палаты
регионов – Совета Федерации. Потому
что именно сенаторы имеют возможность оценивать ситуацию не только в региональном масштабе, но в связке с межрегиональными, федеральными проектами и программами.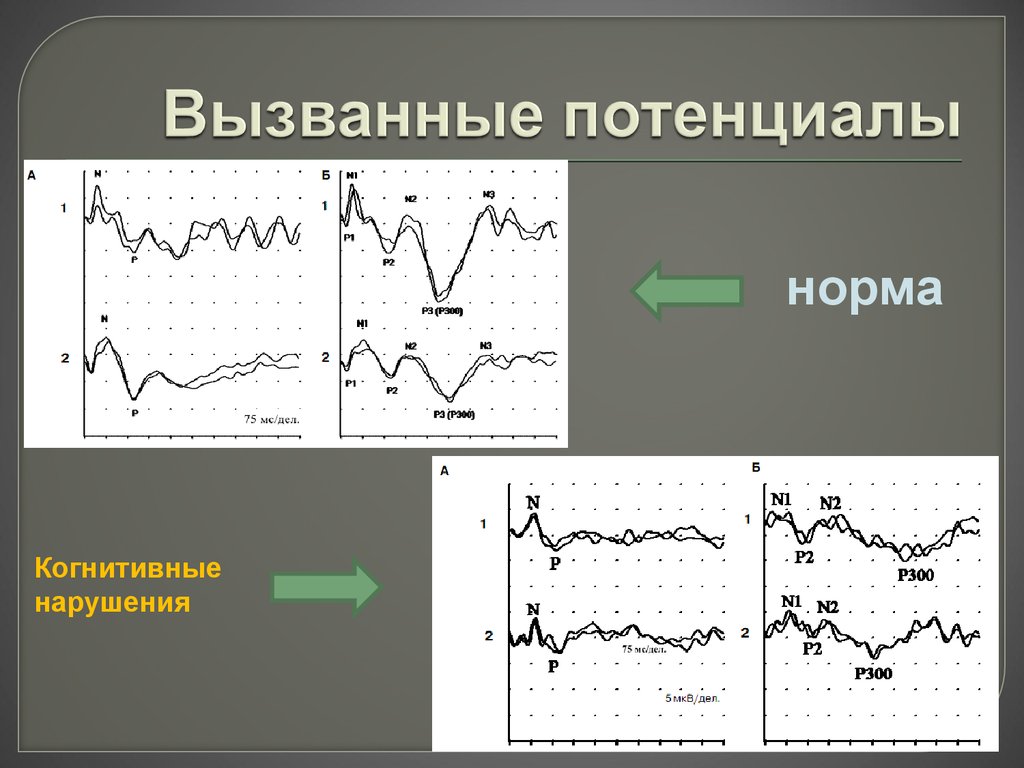 Мы будем держать в поле зрения это направление,
способствовать решению встающих здесь вопросов, проблем. Необходимые для этого
полномочия и инструменты у палаты имеются – от модернизации законодательства до парламентского контроля. И мы намерены их использовать самым активным образом.
Ведь речь идёт о задачах, имеющих ключевое значение не только для отдельных
регионов, а для всей страны.
Мы будем держать в поле зрения это направление,
способствовать решению встающих здесь вопросов, проблем. Необходимые для этого
полномочия и инструменты у палаты имеются – от модернизации законодательства до парламентского контроля. И мы намерены их использовать самым активным образом.
Ведь речь идёт о задачах, имеющих ключевое значение не только для отдельных
регионов, а для всей страны.
Известная истина
гласит, что сильная Россия — это сильные регионы. Поэтому я полностью согласна
с главой Российского государства в том, что у нас не должно быть так называемых
неперспективных регионов, оказавшихся в стороне от экономического роста.
Инвестиционный, экономический потенциал есть у каждого субъекта Федерации.
Необходимо раскрыть, в полной мере реализовать его. В своих выступлениях на площадке Санкт-Петербургского форума Президент В.В. Путин предложил ряд новых
инструментов решения этой задачи. Их
много, но в приоритете направление ресурсов на создание
комфортной среды для жизни людей. Потому что сейчас именно среда обитания
определяет останутся ли люди работать на развитие своего регион, или уедут
туда, где лучше дороги, есть хорошие школы, больницы, а свободное время можно
проводить в парках и музеях.
Потому что сейчас именно среда обитания
определяет останутся ли люди работать на развитие своего регион, или уедут
туда, где лучше дороги, есть хорошие школы, больницы, а свободное время можно
проводить в парках и музеях.
В этой связи,
считаю, назрела необходимость корректировки, а если называть вещи своими
именами, — коренной переработки Стратегии пространственного развития Российской
Федерации на период до 2025 года. Я уже высказывала свою точку зрения по поводу
данного документа. Главный недостаток концепции вижу в том, что она не сбалансирована. Во главу угла поставлен курс на поддержку агломераций,
создаваемых на основе перспективных субъектов Российской Федерации. Так
называемые неперспективные регионы обречены либо на растворение внутри
агломераций, либо на экономическое и социальное умирание. Что в свою очередь
чревато исчезновением населённых пунктов, обезлюживанием значительных
территорий страны, ухудшением экологической ситуации в местах сверхплотной
застройки. И это в условиях и без того неблагоприятной демографической ситуации
в стране.
Повторю, стратегия нуждается в коренной переработке, в основу которой теперь должен быть положен принцип, сформулированный Президентом страны на форуме в Петербурге – в России нет субъектов Федерации, не обладающих потенциалом развития. Считаю, что без учета этой идеи ни законодатели, ни Правительство, ни регионы не имеют права дальше строить свою работу. Тем более, что за три с лишним десятилетия развития новой России мы накопили достаточно опыта и знаний.
Потенциал электрического поля. Разность потенциалов 10 класс онлайн-подготовка на Ростелеком Лицей
Введение
Электрическое поле действует на помещенный в него заряд с силой, которая определяется величиной заряда и напряженностью поля в данной точке.
Если эта сила перемещает заряд – то она совершает работу. Даже если заряда в поле нет, то потенциально эта работа все равно может быть совершена, как только он там окажется. Из опыта других разделов физики мы знаем, что работа связана с энергией.
Для решения некоторых задач удобно использовать энергетическую модель описания электрического поля. Проведем аналогию с гравитационным полем.
Понятие потенциала
Если мы поднимем тело массы , лежащее на земле на высоту (см. рис. 1), мы изменим его потенциальную энергию на величину . Именно такую работу и необходимо совершить для этого подъема.
Рис. 1. Изменение потенциальной энергии
Для любой массы разница энергий на высоте 0 и будет равна (см. рис. 2).
Рис. 2. Разница потенциальных энергий
Если разделить значение потенциальной энергии на массу, мы получим величину, характеризующую гравитационное поле в данной точке. Выражение уже не зависит от массы, оно показывает работу, которую необходимо совершить для переноса тела, с некоторой массой, на высоту , деленную на эту массу.
Теперь посмотрим, как ввести аналог потенциальной энергии приведенной на единицу массы в электрическом поле.
На заряд , находящийся в поле другого заряда , закрепленного в некоторой точке пространства, действует сила Кулона . Эта сила может переместить заряд , совершив при этом работу. Значит, система двух зарядов, находящихся на определенном расстоянии, обладает потенциальной энергией, зависящей от величины зарядов и расстояния между ними.
Если по аналогии с гравитационным полем рассмотреть величину, равную этой энергии, деленной на заряд , то она уже не будет зависеть от заряда и охарактеризует только поле заряда в данной точке. То есть будет являться функцией заряда и расстояния между зарядами. Эта величина и называется потенциалом электрического поля.
Разность потенциалов двух точек, умноженная на величину заряда , равна работе, необходимой для перемещения этого заряда между этими точками. То есть разность потенциалов двух точек поля – это работа по перемещению между ними единичного заряда.
Как и в поле сил тяжести, эта работа не зависит от траектории и определяется только положением точек, между которыми перемещается единичный заряд. Такие поля называют консервативными. В разделе «Механика» мы уже говорили, что энергия – величина, требующая для измерения задания «начала отсчета». Например, в гравитационном поле мы можем считать нулевой потенциальную энергию тела, находящегося на уровне земли. В случае электростатического поля, создаваемого зарядом, естественно считать нулевой потенциальной энергией некоторого заряда, находящегося в поле, его энергию на бесконечном удалении от заряда, в поле которого он находится. Это и есть «точка отсчета» для потенциальной энергии поля заряда.
Такие поля называют консервативными. В разделе «Механика» мы уже говорили, что энергия – величина, требующая для измерения задания «начала отсчета». Например, в гравитационном поле мы можем считать нулевой потенциальную энергию тела, находящегося на уровне земли. В случае электростатического поля, создаваемого зарядом, естественно считать нулевой потенциальной энергией некоторого заряда, находящегося в поле, его энергию на бесконечном удалении от заряда, в поле которого он находится. Это и есть «точка отсчета» для потенциальной энергии поля заряда.
Потенциал поля в некоторой точке равен работе по перемещению единичного заряда из этой точки на бесконечность.
Выражение для потенциала поля точечного заряда
Пусть положительный заряд находится на расстоянии от положительного заряда (см. рис. 3).
Рис. 3. Изначальное положение заряда
Какую работу совершит электрическое поле при перемещении заряда вдоль радиуса в точку, отдаленную на от ? (см. рис. 4)
рис. 4)
Рис. 4. Конечное положение заряда
По определению работа силы равна этой силе, умноженной на перемещение:
В данном случае действует сила электрического взаимодействия (см. рис. 5), по закону Кулона .
Рис. 5. Действие силы электрического взаимодействия
Сила и перемещение в нашем случае сонаправлены, и . Так мы можем находить работу для случая, когда сила постоянна на всей траектории. Здесь же сила изменяется по мере отдаления зарядов друг от друга.
Обозначим перемещение заряда (см. рис. 6).
Рис. 6. Перемещение заряда
По мере перемещения заряда сила изменяется, но на малом (в сравнении с расстоянием до заряда ) отрезке можем считать ее постоянной и находить работу по определению, которое мы привели выше.
Работа, совершаемая силой Кулона на таком малом отрезке равна , где силу можно считать постоянной на всем отрезке . Тогда работа при перемещении на расстояние будет равна сумме работ на участках (), на каждом из которых сила Кулона постоянна и равна .
Эта сумма будет равна
Подробный вывод этой формулы вы можете проследить в ответвлении.
Работа при перемещении электрического заряда
Работа по перемещению заряда на малом участке равна:
Работа на участке равна сумме работ на каждом участке :
Воспользуемся приближенным равенством:
Прежде чем его применить, покажем, что равенство справедливо. Приведем правую часть к общему знаменателю:
Раскроем скобки:
Заметим, что – пренебрежимо малая по сравнению с величина, не может считаться пренебрежимо малой, т. к. количество участков велико. Поэтому в знаменателе можем пренебречь членами и .
Вернемся к нахождению работы. Распишем выражение по полученной формуле:
Распишем сумму:
Мы знаем, что работа связана с энергией. Система обладает энергией, если силы, возникающие в системе, могут выполнить работу (в нашем случае это сила электростатического взаимодействия зарядов). Работа равна уменьшению потенциальной энергии:
Работа равна уменьшению потенциальной энергии:
Сравнив с выражением , делаем вывод, что – это потенциальная энергия взаимодействия двух зарядов. Ранее мы приняли, что потенциальная энергия заряда, отдаленного от источника электрического поля на бесконечность, равна нулю. Посмотрим, как с этим согласуется полученная формула:
Действительно, будет равна нулю на бесконечном отдалении от заряда , т. к. при .
Теперь проверим, как полученный результат соотносится с моделью, в которой разноименные заряды обозначены знаками плюс и минус. Если заряды одноименные, то потенциальная энергия взаимодействия положительна . Система стремится к состоянию с наименьшей потенциальной энергией (как и, например, камень на некоторой высоте над поверхностью земли, предоставленный сам себе, будет падать вниз, т. е. уменьшать высоту и с ней потенциальную энергию )
Действительно, заряды будут отталкиваться и сила электрического взаимодействия будет вызывать перемещение заряда на большее расстояние, потенциальная энергия будет уменьшаться.
Если заряды разноименные, то потенциальная энергия взаимодействия имеет знак минус. Заряды притягиваются, и сила их взаимодействия вызывает перемещение заряда на меньшее расстояние , потенциальная энергия уменьшается.
Потенциал электрического поля
Энергия заряда в поле заряда , равная , зависит от величин обоих зарядов. Характеристика поля, созданного зарядом , естественно, не должна зависеть от величины помещенного в него заряда. Разделим на и получим . Эта величина называется потенциалом электрического поля и обозначается буквой . Эта характеристика поля показывает, какой энергией обладает положительный заряд, помещенный в данную точку поля. Как и энергия, потенциал – скалярная величина, измеряется в вольтах.
В нашем случае – потенциал поля точечного заряда. Точка отсчета потенциалов в нашем случае естественным образом является бесконечно отдаленной точкой (см. рис. 7).
Рис. 7. Точка отсчета потенциалов
В зависимости от задачи точкой отсчета выбирают потенциал поверхности Земли, потенциал отрицательно заряженной пластины конденсатора или потенциал любой другой точки, удобной для решения задачи.
Таким образом, пользуясь определением потенциала, можно вычислить потенциальную энергию заряда, находящегося в электростатическом поле:
и работу поля по перемещению заряда из точки с потенциалом в точку с потенциалом :
Электрическое поле является консервативным, его работа не зависит от траектории движения заряда, а зависит только от перемещения.
Заряд всегда распределен на каком-то теле, имеющем геометрические размеры. На расстояниях, много больших размеров тела, поле слабо зависит от объема и формы этого тела, и потому модели точечного заряда достаточно. Например, потенциал поля заряженного металлического шара при эквивалентен потенциалу поля точечного заряда (см. рис. 8):
Рис. 8. Потенциал поля при
.
Внутри шара потенциал во всех точках одинаков и равен потенциалу на поверхности шара (см. рис. 9):
Рис. 9. Потенциал внутри шара
.
Если бы это было не так, то потенциальная энергия в разных точках внутри шара отличалась бы, а, так как внутри металла есть свободные носители заряда, поле выполняло бы работу по перемещению зарядов. В итоге электроны переместились бы в область большего потенциала, тем самым уменьшив его. Таким образом, потенциал во всех точках приравнивается.
В итоге электроны переместились бы в область большего потенциала, тем самым уменьшив его. Таким образом, потенциал во всех точках приравнивается.
Потенциал подчиняется принципу суперпозиции. При наличии нескольких источников поля складываются как векторы напряженности поля, так и потенциалы:
Задача 1
При перемещении заряда между точками с разностью потенциалов 1 кВ электрическое поле совершило работу 40 мкДж. Чему равен заряд?
Это простая задача на понимание смысла величины разности потенциалов.
Разность потенциалов равна работе по переносу заряда, деленной на величину этого заряда.
Выразим значение заряда:
И вычислим ответ:
Ответ:
Задача 2
Какую работу надо совершить, чтобы перенести заряд 5 мкКл из бесконечности в точку поля, удаленную от центра заряженного шара на 18 см? Заряд шара – 20 мкКл.
Порассуждаем.
- Потенциал поля заряженного шара на бесконечности равен нулю. Следовательно, приближая заряд от бесконечности к шару, внешней силе нужно совершать работу для преодоления силы электростатического взаимодействия. Численно эта работа будет равна работе электрического поля заряженного шара по перемещения заряда с расстояния 18 см на бесконечность.
- Работа по переносу заряда в электрическом поле связана с разностью потенциалов между начальной и конечной точками траектории и величиной заряда
- Величина переносимого заряда у нас есть.
- Потенциал поля заряженного шара на бесконечности, как мы уже отметили, равен нулю. А в конечной точке траектории мы сможем его вычислить, пользуясь формулой для потенциала поля точечного заряда, которая справедлива и для поля вне заряженного шара.
 Следовательно, приближая заряд от бесконечности к шару, внешней силе нужно совершать работу для преодоления силы электростатического взаимодействия. Численно эта работа будет равна работе электрического поля заряженного шара по перемещения заряда с расстояния 18 см на бесконечность.
Следовательно, приближая заряд от бесконечности к шару, внешней силе нужно совершать работу для преодоления силы электростатического взаимодействия. Численно эта работа будет равна работе электрического поля заряженного шара по перемещения заряда с расстояния 18 см на бесконечность.Приступим к решению.
Найдем потенциал электрического поля заряженного шара в конечной точке траектории.
Потенциал электрического поля заряженного шара на бесконечности равен нулю.
Разность потенциалов электрического поля по переносу заряда из точки с потенциалом в точку с потенциалом будет равна:
В то же время она будет равна работе электрического поля по переносу заряда, деленной на заряд:
Величина работы внешних сил, которую надо совершить, чтобы перенести заряд из точки с меньшим потенциалом в точку с большим потенциалом, равна работе электрического поля по переносу такого же заряда в обратном направлении.
Таким образом, мы получили систему из пяти уравнений, решив которую найдем искомую величину. Пронаблюдать математическую часть решения задачи вы можете в свертке.
Ответ: .
Математическая часть решения задачи 2
Подставим выражения для потенциалов из первого и второго уравнений в третье:
Подставим полученную разность потенциалов в четвертое уравнение.
И выразим работу электрического поля:
Согласно пятому уравнению это и есть искомая работа .
Подставим данные из условия и рассчитаем ответ:
Задача решена.
На этом наш урок закончен. Спасибо за внимание.
Список литературы
- Соколович Ю. А., Богданова Г. С. Физика: Справочник с примерами решения задач. – 2-е издание передел. – X.: Веста: Издательство «Ранок», 2005. – 464 с.
- Мякишев Г. Я., Буховцев Б. Б., Сотский Н. Н. Физика: Учеб. для общеобразоват. учреждений. Базовый и профильный уровни. 19-е издание – М.: Просвещение, 2010.
 учреждений. Базовый и профильный уровни. 19-е издание – М.: Просвещение, 2010.
учреждений. Базовый и профильный уровни. 19-е издание – М.: Просвещение, 2010.
Дополнительные рекомендованные ссылки на ресурсы сети Интернет
- Интернет-сайт «phyzika.ru» (Источник)
- Интернет-сайт «physics.ru» (Источник)
- Интернет-сайт «knowlegeport.narod.ru» (Источник)
Домашнее задание
- Какой вид имеет формула для работы электрического поля?
- Что такое потенциал электрического поля?
- Решите задачу: точечный заряд , находясь в некоторой точке поля, обладает потенциальной энергией 1 мкДж. Найдите потенциал этой точки поля.
Не в отметках счастье: что такое личностный потенциал
У каждого человека есть потенциал, и очень важно реализовать его. Но как потенциальная внутренняя энергия становится кинетической? Кажется, что-то подобное нам говорили на уроках физики. Но в жизни всё не так просто. А как именно, давайте разбираться вместе.
Мы привыкли, что учителя, родители, образовательная система в целом и даже сами дети оценивают свои способности как нечто фиксированное и заданное заранее: гуманитарий или технарь, талантливый или бездарный. Математические, социально-экономические классы точно так же делят учеников на тех, у кого «получается» и «не получается». Мы делим детей на одарённых и нет, часто не думая о том, сколько мы сделали, чтобы дать ребёнку возможность раскрыться и показать себя вне шаблона. А современная система оценок и экзаменов только усугубляет ситуацию, оценивая лишь знания и игнорируя особенности личности и потенциала ребёнка.
Что такое личностный потенциал?
Согласно исследованиям, самодисциплина больше влияет на академические успехи, чем IQ. И даже для творчества личность важнее, чем одаренность. Способности важны, но куда важнее умение использовать свои способности — это и называется личностным потенциалом.
Он нужен для решения ключевых вызовов, с которыми человек сталкивается каждый день: принять решение в ситуации неопределенности, достичь поставленной цели, а в ситуации угрозы и давления — сохранить себя и измениться, при этом не изменяя себе. В этом помогают разные компоненты личностного потенциала, в том числе эти три:
В этом помогают разные компоненты личностного потенциала, в том числе эти три:
- Жизнестойкость — способность справляться со стрессовыми ситуациями, сохраняя самообладание. Жизнестойкий человек уверенно принимает решения, берёт на себя ответственность за них и получает удовольствие от ощущения контроля за собственной жизнью. Он осознаёт, что рисков не избежать, и принимает их. А ещё творчески мыслит, умеет договариваться и сотрудничать с другими людьми.
- Мотивационная направленность — то, что движет человеком. Стимулы могут быть как внешними, так и внутренними. К примеру, кому-то важнее получить признание и награду за достижение, а другому достаточно чувства выполненного долга и удовлетворения.
- Оптимизм — не только «стакан заполнен наполовину». Оптимисты куда реже сталкиваются с выученной беспомощностью, им легче налаживать контакты и строить долгосрочные отношения, договариваться с другими. А ещё они ведут более здоровый образ жизни и раньше пессимистов обращаются к врачу.

По результатам исследований российских психологов Тамары Гордеевой, Дмитрия Леонтьева и Евгения Осина, у школьников и студентов, которые ориентированы на состояние потока и получение высоких оценок, целеустремлённы, настойчивы (всё это — элементы личностного потенциала), успехи в учёбе в целом гораздо лучше и заметнее. Особенно если сравнивать их со сверстниками, чей личностный потенциал не так развит.
Тем не менее исследователи обнаружили и неблагополучных студентов, которые достигали неменьших академических высот. При этом их показатели жизнестойкости, целеустремлённости и ощущения осмысленности жизни были ниже, чем у других респондентов.
В качестве примера они приводят ситуацию с участниками школьных олимпиад: те дети, которые стремятся к призовым местам ради внешнего признания и славы, льготного поступления в вуз, оказываются менее успешны в учёбе в целом. Куда более высокий уровень личностного потенциала показывают их сверстники, для которых олимпиада — это интересное испытание, которое ценно само по себе. Стабильно высоких результатов в учёбе обычно добиваются те, кто осознаёт её важность, учится с удовольствием и интересом.
Стабильно высоких результатов в учёбе обычно добиваются те, кто осознаёт её важность, учится с удовольствием и интересом.
«Победители олимпиад высокого уровня сложности отличаются определёнными особенностями мышления — критичностью и независимостью. В отличие от героя Вольтера, они не склонны считать, что „всё к лучшему в этом лучшем из миров“, критически оценивая своё будущее и окружающий мир, школу и жизнь в целом. Они принимают вызовы, бросаемые жизнью, будучи убеждены в том, что всё то, что с ними случается, способствует их развитию за счёт приобретаемого опыта — позитивного или негативного».
Отдельно учёные выделяют оптимистический стиль — подход к жизни, благодаря которому человек оценивает успехи и достижения не как случайное стечение обстоятельств, а как закономерный результат усердной и кропотливой работы. Тем не менее у многих олимпиадников стиль как раз пессимистический. Они ждут худшего исхода, а потому готовятся ко всем возможным (в том числе самым сложным) вариантам заданий.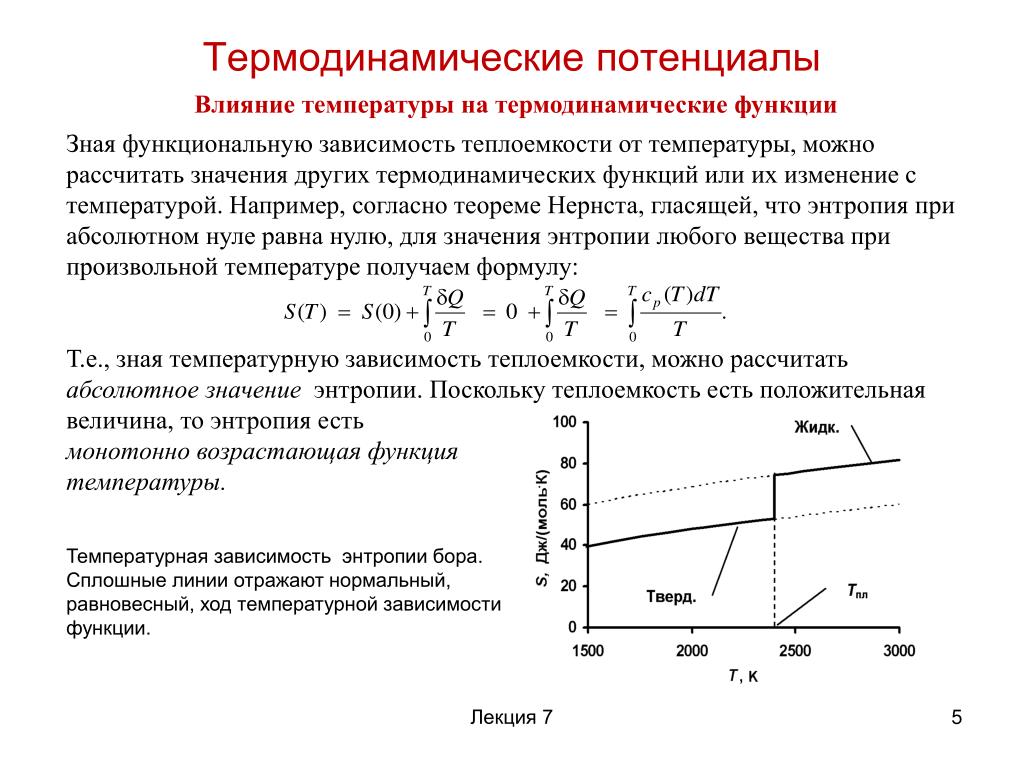
Как же его можно развить
Личностный потенциал состоит из множества компонентов, которые связаны друг с другом. Кроме того, хотя они и развиваются на протяжении всей жизни, многие закладываются в раннем детстве, внутри семьи. И чем раньше родители начнут заниматься с ребёнком, тем лучше.
При этом принудительные занятия арифметикой и чтением, дисциплина и муштра — совсем не главное. Родитель должен видеть в ребёнке личность, возможности которой практически безграничны. Его главная задача — показать те перспективы и направления, в которых ребёнок может развиваться. Ведь составляющие, из которых складывается личностный потенциал, универсальны, то есть их можно применять в любой сфере. Следовательно, и развивать их можно с помощью разных активностей.
«Что такое школа? Один из могучих вечных институтов приобщения человека к культуре человечества. А говоря на скучном языке исследования, один из институтов социализации личности. Это главная характеристика. Для меня на первый план выходит школа, которая помогает в общении с учителями и сверстниками обрести своё „я“. Вещи, связанные со знаниями, умениями и навыками, — второстепенный момент в этом процессе. Школа должна давать картину мира. И сегодняшняя школа должна быть школой неопределённости, школой обучения поведению в сложных ситуациях, школой, учитывающей разнообразие», — говорит об образовании психолог Александр Асмолов.
Вещи, связанные со знаниями, умениями и навыками, — второстепенный момент в этом процессе. Школа должна давать картину мира. И сегодняшняя школа должна быть школой неопределённости, школой обучения поведению в сложных ситуациях, школой, учитывающей разнообразие», — говорит об образовании психолог Александр Асмолов.
Ещё в начале прошлого века советский психолог Лев Выготский подчёркивал важность окружающей среды для гармоничного развития ребёнка. Он утверждал, что человек формирует высшие психические функции, приобретает навыки и знания в тот момент, когда переносит их из внешнего мира во внутренний. Этот процесс он называл интериоризацией (да, мы понимаем, сложное слово, но за ним стоит важное понятие). Продолжая традицию легендарного психолога, современные методисты и педагоги разработали концепцию личностно-развивающей среды.
Её суть в том, что ребёнок живёт и учится в условиях, где всё создано для самостоятельного обучения и саморазвития. Здесь каждая деталь играет свою роль: учебные классы, школьный двор, игровые площадки, проблемно-ориентированный подход к преподаванию, доверительные и открытые отношения с учителями, — важно всё.
Находясь в такой среде, детям комфортно общаться друг с другом и со взрослыми, они охотно проявляют инициативу и учатся быть проактивными, ставят перед собой волнующие их вопросы и ищут на них ответы. Дружественная атмосфера позволяет каждому ребёнку спокойно высказывать своё мнение, делиться желаниями, страхами, планами и мечтами.
Такая школа идёт по пути развития навыков в комплексе с достижением личностных результатов. Причём очень важно, чтобы эта среда начиналась не с первого класса, а ещё с дошкольного этапа. И чтобы в ней вместе с детьми на равных работали и родители, и педагоги. Создание такой среды — это первый шаг к созданию школы возможностей, школы, в которой развивается личностный потенциал ребенка.
При подготовке текста мы использовали материалы программы БФ Сбербанка «Вклад в будущее» по развитию личностного потенциала.
Потенциалы покоя и потенциалы действия (Раздел 1, Глава 1) Неврология в Интернете: Электронный учебник по неврологии | Кафедра нейробиологии и анатомии
 youtube.com/embed/ZY6CL2D_SpA?rel=0″ frameborder=»0″ allowfullscreen=»»> youtube.com/embed/ZY6CL2D_SpA?rel=0″ frameborder=»0″ allowfullscreen=»»> |
| Видео лекции |
Несмотря на огромную сложность мозга, можно получить представление о его функциях, обратив внимание на две основные детали:
- Во-первых, способы, которыми отдельные нейроны, составляющие нервную систему, соединяются вместе, чтобы генерировать поведение.
- Во-вторых, биофизические, биохимические и электрофизиологические свойства отдельных нейронов.
Лучше всего начать с компонентов нервной системы и того, как электрические свойства нейронов наделяют нервные клетки способностью обрабатывать и передавать информацию.
| Видео лекции |
1.1 Введение в потенциал действия
Рисунок 1. |
 1
1 Теории кодирования и передачи информации в нервной системе восходят к греческому врачу Галену (129-210 г. н.э.), который предложил гидравлический механизм, с помощью которого мышцы сокращаются из-за поступления в них жидкости из полых нервов. Эта базовая теория существовала веками и была развита Рене Декартом (1596–1650), который предположил, что духи животных вытекают из мозга через нервы, а затем в мышцы, чтобы производить движения (см. эту анимацию для современной интерпретации такой гидравлической теории нервных функция). Серьезный сдвиг парадигмы произошел с новаторской работой Луиджи Гальвани, который обнаружил в 179 г.4, что нерв и мышца могут быть активированы заряженными электродами, и предположил, что нервная система функционирует посредством передачи электрических сигналов (см. анимацию эксперимента Гальвани). Однако среди ученых возникли споры о том, находится ли электричество в нервах и мышцах или нервы и мышцы просто реагируют на вредный электрический удар через какой-то внутренний неэлектрический механизм. Проблема не была решена до 1930-х годов с разработкой современных электронных усилителей и записывающих устройств, позволяющих записывать электрические сигналы. Одним из примеров является новаторская работа Х.К. Hartline 80 лет назад по электрической сигнализации мечехвоста Limulus. Электроды располагали на поверхности зрительного нерва. (Помещая электроды на поверхность нерва, можно получить представление об изменениях мембранного потенциала, происходящих между внешней и внутренней частью нервной клетки.) Затем производились вспышки света различной интенсивности длительностью 1 с. предстал перед глазами; сначала тусклый свет, затем более яркий свет. Очень тусклый свет не вызывал изменений в активности, но более яркий свет вызывал небольшие повторяющиеся пикообразные события. Эти спайкоподобные события называются потенциалами действия, нервными импульсами или иногда просто спайками. Потенциалы действия — это основные события, которые нервные клетки используют для передачи информации из одного места в другое.
Проблема не была решена до 1930-х годов с разработкой современных электронных усилителей и записывающих устройств, позволяющих записывать электрические сигналы. Одним из примеров является новаторская работа Х.К. Hartline 80 лет назад по электрической сигнализации мечехвоста Limulus. Электроды располагали на поверхности зрительного нерва. (Помещая электроды на поверхность нерва, можно получить представление об изменениях мембранного потенциала, происходящих между внешней и внутренней частью нервной клетки.) Затем производились вспышки света различной интенсивности длительностью 1 с. предстал перед глазами; сначала тусклый свет, затем более яркий свет. Очень тусклый свет не вызывал изменений в активности, но более яркий свет вызывал небольшие повторяющиеся пикообразные события. Эти спайкоподобные события называются потенциалами действия, нервными импульсами или иногда просто спайками. Потенциалы действия — это основные события, которые нервные клетки используют для передачи информации из одного места в другое.
1.2 Особенности потенциалов действия
Записи на рисунке выше иллюстрируют три очень важные особенности нервных потенциалов действия. Сначала потенциал действия нерва имеет короткую продолжительность (около 1 мс). Второй , нервные потенциалы действия вызываются по принципу «все или ничего». Третий , нервные клетки кодируют интенсивность информации частотой потенциалов действия. При увеличении интенсивности раздражителя размер потенциала действия не увеличивается. Скорее увеличивается частота или количество потенциалов действия. Как правило, чем выше интенсивность раздражителя (будь то световой раздражитель фоторецептора, механический раздражитель кожи или растяжение мышечного рецептора), тем большее количество вызванных потенциалов действия. Точно так же для двигательной системы: чем больше потенциалов действия в мотонейроне, тем больше интенсивность сокращения мышцы, иннервируемой этим мотонейроном.
Потенциалы действия имеют большое значение для функционирования мозга, поскольку они передают информацию из нервной системы в центральную нервную систему и передают команды, инициированные в центральной нервной системе, на периферию. Следовательно, необходимо досконально разобраться в их свойствах. Чтобы ответить на вопросы о том, как инициируются и распространяются потенциалы действия, нам необходимо зарегистрировать потенциал между внутренней и внешней частью нервных клеток, используя методы внутриклеточной записи.
Следовательно, необходимо досконально разобраться в их свойствах. Чтобы ответить на вопросы о том, как инициируются и распространяются потенциалы действия, нам необходимо зарегистрировать потенциал между внутренней и внешней частью нервных клеток, используя методы внутриклеточной записи.
1.3 Внутриклеточные записи нейронов
Рисунок 1.2 |
Разность потенциалов на мембране нервной клетки можно измерить с помощью микроэлектрода , наконечник которого настолько мал (около микрона), что он может проникать в клетку, не вызывая повреждения. Когда электрод находится в ванне (внеклеточной среде), потенциал не регистрируется, поскольку ванна изопотенциальна. Если микроэлектрод осторожно ввести в ячейку, происходит резкое изменение потенциала. Показание вольтметра мгновенно изменяется от 0 мВ до показания разности потенциалов -60 мВ внутри ячейки по отношению к внешней стороне. Потенциал, который регистрируется, когда живую клетку пронзают микроэлектродом, называется потенциалом покоя и варьируется от клетки к клетке. Здесь показано, что оно составляет -60 мВ, но может варьироваться от -80 мВ до -40 мВ, в зависимости от конкретного типа нервной клетки. В отсутствие какой-либо стимуляции потенциал покоя обычно постоянен.
Потенциал, который регистрируется, когда живую клетку пронзают микроэлектродом, называется потенциалом покоя и варьируется от клетки к клетке. Здесь показано, что оно составляет -60 мВ, но может варьироваться от -80 мВ до -40 мВ, в зависимости от конкретного типа нервной клетки. В отсутствие какой-либо стимуляции потенциал покоя обычно постоянен.
Также можно записывать и изучать потенциал действия. На рис. 1.3 показан пример, в котором нейрон уже проколот одним микроэлектродом (записывающим электродом), который подключен к вольтметру. Электрод регистрирует потенциал покоя -60 мВ. В клетку также был проколот второй электрод, называемый стимулирующим электродом. Этот электрод подключен к батарее и устройству, которое может контролировать величину тока (I), протекающего через электрод. Изменения мембранного потенциала вызываются замыканием переключателя и систематическим изменением размера и полярности батареи. Если отрицательный полюс батареи соединить с внутренней частью ячейки, как показано на рис. 1.3А, через стимулирующий электрод будет протекать мгновенное изменение величины тока, и мембранный потенциал временно станет более отрицательным. Этот результат не должен вызывать удивления. Отрицательный полюс батареи делает внутреннюю часть ячейки более отрицательной, чем она была раньше. Изменение потенциала, увеличивающее поляризованное состояние мембраны, называется гиперполяризация . Клетка более поляризована, чем обычно. Используйте еще большую батарею, и потенциал станет еще больше. Результирующие гиперполяризации являются градуированными функциями величины стимулов, использованных для их создания.
1.3А, через стимулирующий электрод будет протекать мгновенное изменение величины тока, и мембранный потенциал временно станет более отрицательным. Этот результат не должен вызывать удивления. Отрицательный полюс батареи делает внутреннюю часть ячейки более отрицательной, чем она была раньше. Изменение потенциала, увеличивающее поляризованное состояние мембраны, называется гиперполяризация . Клетка более поляризована, чем обычно. Используйте еще большую батарею, и потенциал станет еще больше. Результирующие гиперполяризации являются градуированными функциями величины стимулов, использованных для их создания.
Рисунок 1.3 |
Теперь рассмотрим случай, когда положительный полюс батареи соединен с электродом (рис. 1.3Б). Когда положительный полюс батареи соединен с электродом, потенциал ячейки становится более положительным, когда переключатель замкнут (рис. 1.3В). Такие потенциалы называются деполяризации . Поляризованное состояние мембраны снижается. Батареи большего размера вызывают еще большую деполяризацию. Опять же, величина ответов пропорциональна величине стимулов. Однако происходит необычное событие, когда величина деполяризации достигает уровня мембранного потенциала, называемого порогом . Инициируется совершенно новый тип сигнала; потенциал действия. Обратите внимание, что если размер батареи увеличить еще больше, амплитуда потенциала действия будет такой же, как и предыдущая (рис. 1.3Б). Процесс возбуждения потенциала действия в нервной клетке аналогичен воспламенению фитиля от источника тепла. Необходима определенная минимальная температура (порог). Температуры ниже порога не приводят к воспламенению предохранителя. Температуры, превышающие пороговую, воспламеняют фитиль точно так же, как и пороговая температура, и фитиль не горит ярче или горячее.
1.3В). Такие потенциалы называются деполяризации . Поляризованное состояние мембраны снижается. Батареи большего размера вызывают еще большую деполяризацию. Опять же, величина ответов пропорциональна величине стимулов. Однако происходит необычное событие, когда величина деполяризации достигает уровня мембранного потенциала, называемого порогом . Инициируется совершенно новый тип сигнала; потенциал действия. Обратите внимание, что если размер батареи увеличить еще больше, амплитуда потенциала действия будет такой же, как и предыдущая (рис. 1.3Б). Процесс возбуждения потенциала действия в нервной клетке аналогичен воспламенению фитиля от источника тепла. Необходима определенная минимальная температура (порог). Температуры ниже порога не приводят к воспламенению предохранителя. Температуры, превышающие пороговую, воспламеняют фитиль точно так же, как и пороговая температура, и фитиль не горит ярче или горячее.
Однако, если надпороговый текущий стимул будет достаточно продолжительным, будет вызвана последовательность потенциалов действия. Как правило, потенциалы действия будут продолжать срабатывать до тех пор, пока действует стимул, причем частота возбуждения пропорциональна величине стимула (рис. 1.4).
Как правило, потенциалы действия будут продолжать срабатывать до тех пор, пока действует стимул, причем частота возбуждения пропорциональна величине стимула (рис. 1.4).
Рисунок 1.4 |
Потенциалы действия не только инициируются по принципу «все или ничего», но и распространяются по принципу «все или ничего». Потенциал действия, инициированный в теле клетки двигательного нейрона в спинном мозге, будет распространяться недекрементным образом до синаптических окончаний этого двигательного нейрона. Опять же, ситуация аналогична горящему предохранителю. Как только фитиль загорится, пламя распространится до его конца.
1.4 Компоненты потенциалов действия
Потенциал действия состоит из нескольких компонентов (рис. 1.3Б). Порог — это значение мембранного потенциала, при достижении которого происходит инициация потенциала действия по принципу «все или ничего». Начальная или восходящая фаза потенциала действия называется деполяризующей фазой или подъемом . Область потенциала действия между уровнем 0 мВ и амплитудой пика составляет перерегулирование . Возврат мембранного потенциала к потенциалу покоя называется фазой реполяризации . Существует также фаза потенциала действия, в течение которой мембранный потенциал может быть более отрицательным, чем потенциал покоя. Эта фаза потенциала действия называется недоразвитием или гиперполяризующим послепотенциалом . На рисунке 1.4 отрицательные импульсы потенциалов действия не становятся более отрицательными, чем потенциал покоя, потому что они «оседлают» постоянный деполяризующий стимул.
Начальная или восходящая фаза потенциала действия называется деполяризующей фазой или подъемом . Область потенциала действия между уровнем 0 мВ и амплитудой пика составляет перерегулирование . Возврат мембранного потенциала к потенциалу покоя называется фазой реполяризации . Существует также фаза потенциала действия, в течение которой мембранный потенциал может быть более отрицательным, чем потенциал покоя. Эта фаза потенциала действия называется недоразвитием или гиперполяризующим послепотенциалом . На рисунке 1.4 отрицательные импульсы потенциалов действия не становятся более отрицательными, чем потенциал покоя, потому что они «оседлают» постоянный деполяризующий стимул.
1.5 Ионные механизмы потенциалов покоя
Перед изучением ионных механизмов потенциалов действия необходимо сначала понять ионные механизмы потенциала покоя. Эти два явления тесно связаны. История потенциала покоя восходит к началу 1900-х годов, когда Юлиус Бернштейн предположил, что потенциал покоя (V m ) равен равновесному потенциалу калия (E K ). Где
История потенциала покоя восходит к началу 1900-х годов, когда Юлиус Бернштейн предположил, что потенциал покоя (V m ) равен равновесному потенциалу калия (E K ). Где
Ключом к пониманию потенциала покоя является тот факт, что ионы неравномерно распределены внутри и снаружи клеток и что клеточные мембраны избирательно проницаемы для разных ионов. К + особенно важен для потенциала покоя. Мембрана обладает высокой проницаемостью для K + . Кроме того, внутри клетки высокая концентрация K + ([K + ] i ), а снаружи клетки низкая концентрация K + ([K + ] или ). Таким образом, K + будет естественным образом перемещаться путем диффузии из области высокой концентрации в область низкой концентрации. Следовательно, положительный K + ионов, покидая внутреннюю поверхность мембраны, оставляют после себя некоторое количество отрицательно заряженных ионов. Этот отрицательный заряд притягивает положительный заряд уходящего иона K + и имеет тенденцию «притягивать его обратно». Таким образом, будет иметь место электрическая сила, направленная внутрь, которая будет стремиться уравновесить диффузионную силу, направленную наружу. В конце концов установится равновесие; сила концентрации, перемещающая K + наружу, уравновешивает электрическую силу, удерживающую его. Потенциал, при котором достигается это равновесие, называется Потенциал равновесия Нернста .
Этот отрицательный заряд притягивает положительный заряд уходящего иона K + и имеет тенденцию «притягивать его обратно». Таким образом, будет иметь место электрическая сила, направленная внутрь, которая будет стремиться уравновесить диффузионную силу, направленную наружу. В конце концов установится равновесие; сила концентрации, перемещающая K + наружу, уравновешивает электрическую силу, удерживающую его. Потенциал, при котором достигается это равновесие, называется Потенциал равновесия Нернста .
Рисунок 1.5 |
Слева показан эксперимент по проверке гипотезы Бернштейна о том, что мембранный потенциал равен потенциалу равновесия Нернста (т. е. V м = E K ).
Концентрацию K + вне клетки систематически варьировали при измерении мембранного потенциала. Также показана линия, предсказываемая уравнением Нернста. Очень близко к этой линии находятся экспериментально измеренные точки. Более того, из-за логарифмической зависимости в уравнении Нернста изменение концентрации K + с коэффициентом 10 приводит к изменению потенциала на 60 мВ.
Очень близко к этой линии находятся экспериментально измеренные точки. Более того, из-за логарифмической зависимости в уравнении Нернста изменение концентрации K + с коэффициентом 10 приводит к изменению потенциала на 60 мВ.
Обратите внимание, однако, что на рисунке слева есть некоторые отклонения от того, что предсказывает уравнение Нернста. Таким образом, нельзя заключить, что V m = E K . Такие отклонения указывают на то, что в генерации потенциала покоя участвует и другой ион. Этот ион Na + . Высокая концентрация Na + вне клетки и относительно низкая концентрация внутри клетки приводит к химической (диффузионной) движущей силе для Na + наплыв. Существует также электрическая движущая сила, поскольку внутренняя часть клетки является отрицательной, и эта отрицательность притягивает положительные ионы натрия. Следовательно, если клетка имеет небольшую проницаемость для натрия, Na + будет перемещаться через мембрану, и мембранный потенциал будет более деполяризованным, чем можно было бы ожидать от равновесного потенциала K + .
1.6 Уравнение Гольдмана-Ходжкина и Катца (GHK)
Когда мембрана проницаема для двух разных ионов, уравнение Нернста больше нельзя использовать для точного определения мембранного потенциала. Однако можно применить уравнение GHK. Это уравнение описывает потенциал через мембрану, которая проницаема как для Na + и К +.
Обратите внимание, что α представляет собой отношение проницаемости Na + (P Na ) к проницаемости K + (P K ). Отметим также, что если проницаемость мембраны для Na + равна 0, то альфа в GHK равна 0, и уравнение Гольдмана-Ходжкина-Каца сводится к равновесному потенциалу Нернста для K + . Если проницаемость мембраны для Na + очень высока, а проницаемость для калия очень низка, [Na 9Члены 0172 + ] становятся очень большими, доминирующими в уравнении по сравнению с членами [K + ], и уравнение ГХК сводится к равновесному потенциалу Нернста для Na + .
Если применить уравнение GHK к тем же данным, что и на рис. 1.5, результат будет намного лучше. Значение альфа, необходимое для получения такой хорошей подгонки, составляло 0,01. Это означает, что проницаемость калия K + в 100 раз превышает проницаемость Na + . Таким образом, потенциал покоя связан не только с высокой проницаемостью для K + . Существует также небольшая проницаемость для Na + , что делает мембранный потенциал немного более положительным, чем он был бы, если бы мембрана была проницаема только для K + .
| |
Рисунок 1.6 |
1.7 Лаборатория мембранного потенциала
Нажмите здесь, чтобы перейти в интерактивную Лабораторию мембранного потенциала и поэкспериментировать с эффектами изменения внешней или внутренней концентрации ионов калия и проницаемости мембран для ионов натрия и калия. Прогнозы делаются с использованием уравнений Нернста и Гольдмана, Ходжкина, Каца.
Прогнозы делаются с использованием уравнений Нернста и Гольдмана, Ходжкина, Каца.
| Лаборатория мембранного потенциала |
Проверьте свои знания
Если мембрана нерва внезапно стала одинаково проницаемой для обоих Na + и K + мембранный потенциал будет:
Если мембрана нерва внезапно стала одинаково проницаемой для обоих Na + и K + мембранный потенциал будет:
Если нервная мембрана вдруг станет одинаково проницаемой как для Na + , так и для K + , мембранный потенциал будет:
Если нервная мембрана вдруг станет одинаково проницаемой как для Na + , так и для K + , мембранный потенциал будет:
Если нервная мембрана вдруг станет одинаково проницаемой как для Na + , так и для K + , мембранный потенциал будет:
Если нервная мембрана вдруг станет одинаково проницаемой как для Na + , так и для K + , мембранный потенциал будет:
При изменении концентрации К + в цитоплазме аксона беспозвоночного до нового значения 200 мМ (Примечание: для этого аксона нормальный [K] o = 20 мМ и нормальный [K] i = 400 мМ):
Если концентрацию К + в цитоплазме аксона беспозвоночного изменить на новое значение 200 мМ (Примечание: для этого аксона нормальный [K] o = 20 мМ и нормальный [K] i = 400 мМ):
Если концентрацию К + в цитоплазме аксона беспозвоночного изменить на новое значение 200 мМ (Примечание: для этого аксона нормальный [K] o = 20 мМ и нормальный [K] i = 400 мМ):
Если концентрацию К + в цитоплазме аксона беспозвоночного изменить на новое значение 200 мМ (Примечание: для этого аксона в норме [K] o = 20 мМ и нормальный [K] i = 400 мМ):
Если концентрацию К + в цитоплазме аксона беспозвоночного изменить на новое значение 200 мМ (Примечание: для этого аксона нормальный [K] o = 20 мМ и нормальный [K] i = 400 мМ):
Если концентрацию К + в цитоплазме аксона беспозвоночного изменить на новое значение 200 мМ (Примечание: для этого аксона в норме [K] o = 20 мМ и нормальный [K] i = 400 мМ):
|
 Приближение к постоянному значению около +55 мВ
Приближение к постоянному значению около +55 мВ Без изменений
Без изменений  Подойдите к новому равновесному потенциалу Na + . Этот ответ НЕВЕРЕН.
Подойдите к новому равновесному потенциалу Na + . Этот ответ НЕВЕРЕН. Приблизьтесь к значению около 0 мВ. Это ПРАВИЛЬНЫЙ ответ!
Приблизьтесь к значению около 0 мВ. Это ПРАВИЛЬНЫЙ ответ! Альфа в уравнении Голдмана должна приближаться к очень высокому значению.
Альфа в уравнении Голдмана должна приближаться к очень высокому значению. Равновесный потенциал K + будет около -60 мВ
Равновесный потенциал K + будет около -60 мВ Поскольку мембранный потенциал в норме составляет -60 мВ и в значительной степени зависит от Е K изменение концентрации калия и, следовательно, E K сделало бы мембранный потенциал более положительным, n или более отрицательным.
Поскольку мембранный потенциал в норме составляет -60 мВ и в значительной степени зависит от Е K изменение концентрации калия и, следовательно, E K сделало бы мембранный потенциал более положительным, n или более отрицательным. Равновесный потенциал K + изменится на 60 мВ. Этот ответ НЕВЕРЕН.
Равновесный потенциал K + изменится на 60 мВ. Этот ответ НЕВЕРЕН.  Мембранный потенциал стал бы больше
отрицательный
Мембранный потенциал стал бы больше
отрицательный Равновесный потенциал K + будет около -60 мВ
Равновесный потенциал K + будет около -60 мВ Равновесный потенциал K + будет около -60 мВ
Равновесный потенциал K + будет около -60 мВНейронаука для детей — потенциал действия
Описание на этой странице как работают нейроны. Я надеюсь, что это объяснение не станет слишком сложным, но важно понимать, как нейроны делают то, что они делают. Есть много деталей, но идите медленно и смотрите на цифры.
Многое из того, что мы
знания о том, как работают нейроны, получены из экспериментов на гигантском аксоне
кальмар. Этот гигантский аксон тянется от головы к хвосту кальмара.
и используется, чтобы помочь кальмару двигаться. Насколько велик этот аксон? Может быть
до 1 мм в диаметре — легко увидеть невооруженным глазом.
Насколько велик этот аксон? Может быть
до 1 мм в диаметре — легко увидеть невооруженным глазом.
Нейроны посылают сообщения электрохимически . Это означает химические вещества вызывают электрический сигнал. Химические вещества в организме являются «электрически заряженные» — когда у них есть электрический заряд, они позвонил ионов. Важными ионами в нервной системе являются натрий и калий (оба имеют 1 положительный заряд, +), кальций (имеет 2 положительный заряд, ++) и хлорид (имеет отрицательный заряд, -). Есть также некоторые отрицательно заряженные белковые молекулы. Также важно помните, что нервные клетки окружены мембраной, которая позволяет некоторым прохождение ионов и блокирует прохождение других ионов. Этот тип мембрана называется полупроницаемой .
Мембранный потенциал покоя
Когда нейрон не
посылая сигнал, он находится «в покое». Когда нейрон находится в состоянии покоя, его внутренняя
нейрона отрицательна по отношению к внешней стороне. Хотя
концентрации различных ионов пытаются сбалансировать с обеих сторон
мембраны, они не могут, потому что клеточная мембрана допускает лишь некоторые
ионы проходят через каналы (ионные каналы). В покое ионы калия
(K + ) могут легко проникать через мембрану. Также в покое,
ионы хлорида (Cl — ) и ионы натрия (Na + ) есть еще
трудный временной переход. Отрицательно заряженные белковые молекулы
(A — ) внутри нейрона не может пересекать мембрану. Помимо этих селективных ионов
каналов, есть насос , который использует энергию для перемещения трех
ионов натрия из нейрона на каждые два введенных иона калия.
Наконец, когда все эти силы уравновешиваются и разница в
измеряется напряжение между внутренней и внешней частью нейрона, у вас есть
потенциал покоя . Мембрана покоя
потенциал нейрона составляет около -70 мВ (мВ=милливольт) — это означает, что
внутри нейрона на 70 мВ меньше, чем снаружи. В покое имеются
относительно больше ионов натрия вне нейрона и больше ионов калия
внутри этого нейрона.
Хотя
концентрации различных ионов пытаются сбалансировать с обеих сторон
мембраны, они не могут, потому что клеточная мембрана допускает лишь некоторые
ионы проходят через каналы (ионные каналы). В покое ионы калия
(K + ) могут легко проникать через мембрану. Также в покое,
ионы хлорида (Cl — ) и ионы натрия (Na + ) есть еще
трудный временной переход. Отрицательно заряженные белковые молекулы
(A — ) внутри нейрона не может пересекать мембрану. Помимо этих селективных ионов
каналов, есть насос , который использует энергию для перемещения трех
ионов натрия из нейрона на каждые два введенных иона калия.
Наконец, когда все эти силы уравновешиваются и разница в
измеряется напряжение между внутренней и внешней частью нейрона, у вас есть
потенциал покоя . Мембрана покоя
потенциал нейрона составляет около -70 мВ (мВ=милливольт) — это означает, что
внутри нейрона на 70 мВ меньше, чем снаружи. В покое имеются
относительно больше ионов натрия вне нейрона и больше ионов калия
внутри этого нейрона.
Потенциал действия
потенциал покоя говорит о том, что происходит, когда нейрон находится в состоянии покоя.
Потенциал действия возникает, когда нейрон
посылает информацию вниз по аксону от тела клетки.
Нейробиологи используют другие слова, такие как «спайк» или «импульс» для обозначения
потенциал действия. Потенциал действия – это взрыв
электрическая активность, создаваемая деполяризующий
текущий . Это означает, что некоторое событие (стимул) вызывает
потенциал покоя приближается к 0 мВ. Когда деполяризация достигает
около -55 мВ нейрон вызовет потенциал действия. Это порог . Если нейрон не достигает этого критического
пороговый уровень, то потенциал действия не сработает. Также, когда
достигается пороговый уровень, возникает потенциал действия фиксированной величины.
всегда срабатывает … для любого данного нейрона размер потенциала действия равен
всегда одно и то же. В одном нерве нет ни больших, ни малых потенциалов действия.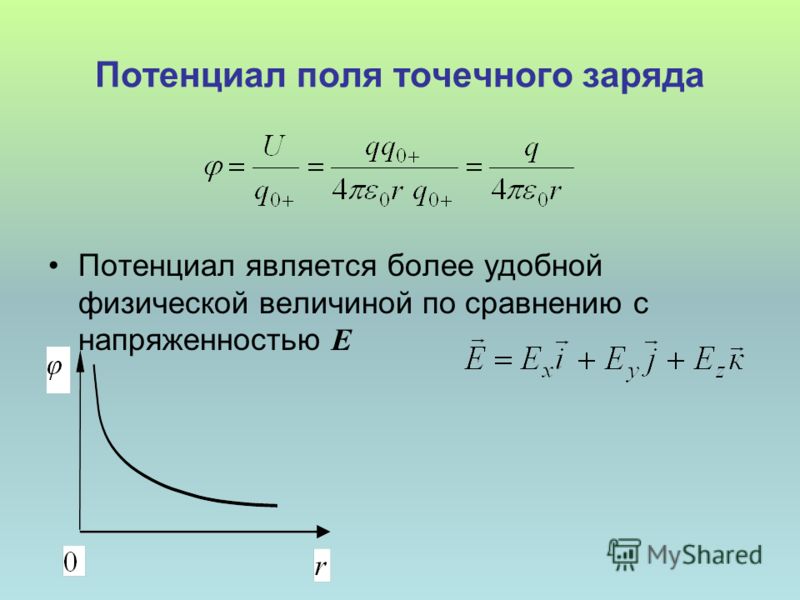 ячейка — все потенциалы действия имеют одинаковую величину. Следовательно, нейрон
либо не достигает порога, либо имеет место полный потенциал действия.
уволен — это принцип «ВСЕ ИЛИ НИКТО».
ячейка — все потенциалы действия имеют одинаковую величину. Следовательно, нейрон
либо не достигает порога, либо имеет место полный потенциал действия.
уволен — это принцип «ВСЕ ИЛИ НИКТО».
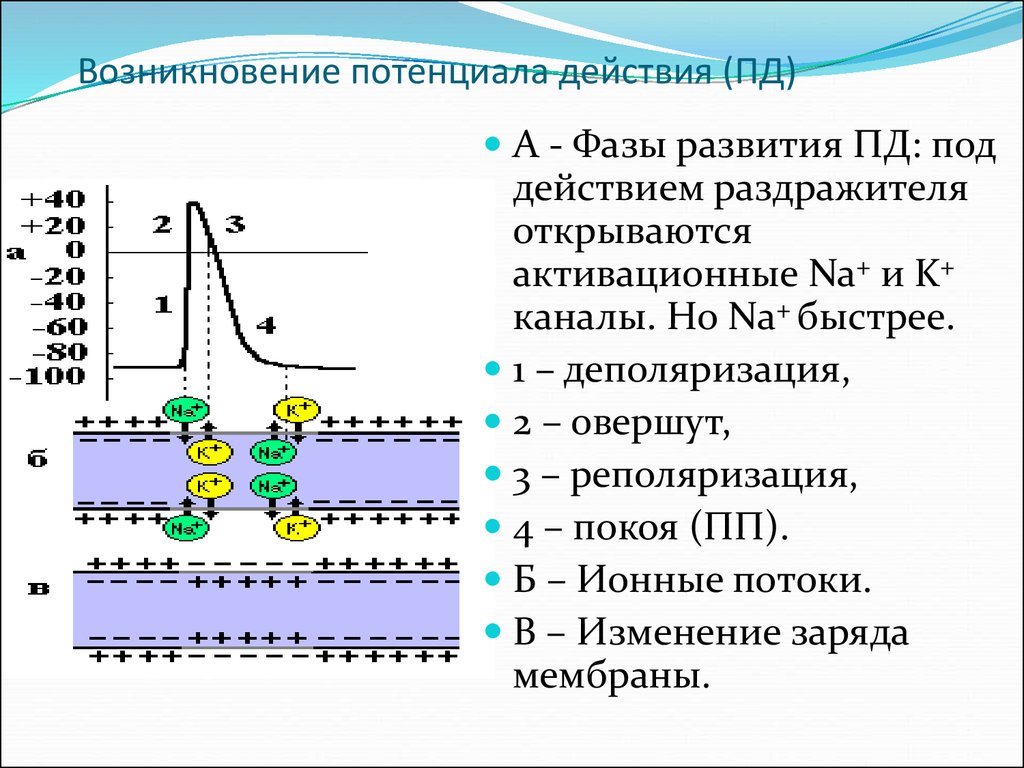
Потенциалы действия вызываются
когда различные ионы пересекают мембрану нейрона. Раздражитель сначала вызывает
натриевые каналы открываются. Поскольку ионов натрия на
снаружи, а внутренняя часть нейрона отрицательна по отношению к внешней,
ионы натрия устремляются в нейрон. Помните, натрий имеет положительный заряд,
поэтому нейрон становится более позитивным и деполяризуется. Занимает
дольше открываются калиевые каналы. Когда они открываются, калий
устремляется из клетки, обращая деполяризацию. Также об этом
время натриевые каналы начинают закрываться. Это приводит к тому, что потенциал действия
вернуться к -70 мВ (реполяризация). Потенциал действия на самом деле
проходит мимо -70 мВ (гиперполяризация), потому что калиевые каналы остаются
открыть слишком долго. Постепенно концентрации ионов возвращаются к состоянию покоя. уровни, и ячейка возвращается к -70 мВ.
уровни, и ячейка возвращается к -70 мВ.
Вот и все… Потенциал действия
| Попробуйте! |
|
| Знаете ли вы? | Гигантский аксон кальмара может быть от 100 до 1000 раз крупнее аксона млекопитающих. Гигантский аксон иннервирует мышца мантии кальмара. Эти мышцы используются для продвижения кальмара через вода. |
Copyright © 1996-2020, Эрик Х. Чудлер Все права Сдержанный.
Потенциалы действия и синапсы — Институт мозга Квинсленда
Ключевые факты: потенциал действия и синапсы
- Нейроны общаются друг с другом посредством электрических событий, называемых «потенциалами действия», и химических нейротрансмиттеров.
- В месте соединения двух нейронов (синапсе) потенциал действия заставляет нейрон А высвобождать химический нейротрансмиттер.
- Нейротрансмиттер может либо помогать (возбуждать), либо препятствовать (ингибировать) нейрону B в возбуждении собственного потенциала действия.
- В интактном мозге баланс сотен возбуждающих и тормозных импульсов нейрона определяет, возникнет ли в результате потенциал действия.

Нейроны — это электрические устройства. В клеточной мембране (граница между внутренней и внешней частью клетки) расположено множество каналов, которые позволяют положительным или отрицательным ионам проникать в клетку и выходить из нее.
В норме внутренняя часть клетки более негативна, чем внешняя; нейробиологи говорят, что внутренняя часть составляет около -70 мВ по отношению к внешней, или что мембранный потенциал клетки в состоянии покоя составляет -70 мВ.
Этот мембранный потенциал не является статическим. Он постоянно поднимается и опускается, в основном в зависимости от сигналов, поступающих от аксонов других нейронов. Некоторые входы заставляют мембранный потенциал нейрона становиться более положительным (или менее отрицательным, например, с -70 мВ до -65 мВ), а другие наоборот.
Некоторые входы заставляют мембранный потенциал нейрона становиться более положительным (или менее отрицательным, например, с -70 мВ до -65 мВ), а другие наоборот.
Их соответственно называют возбуждающими и тормозными входами, поскольку они стимулируют или подавляют генерацию потенциалов действия (причина, по которой одни входы являются возбуждающими, а другие — тормозными, заключается в том, что разные типы нейронов выделяют разные нейротрансмиттеры; нейротрансмиттер, используемый нейроном, определяет его эффект).
Потенциалы действия являются основными единицами связи между нейронами и возникают, когда сумма всех возбуждающих и тормозных импульсов заставляет мембранный потенциал нейрона достигать примерно -50 мВ (см. диаграмму), значение, называемое порог потенциала действия .
Нейробиологи часто называют потенциалы действия «спайками» или говорят, что нейрон «выстрелил спайком» или «спайком». Этот термин относится к форме потенциала действия, регистрируемой с помощью чувствительного электрического оборудования.
Нейрон всплеск, когда комбинация всех полученных им возбуждения и торможения достигает порога. Справа пример реального нейрона коры головного мозга мыши. (Изображение: Алан Вудрафф / QBI)
Синапсы: как нейроны общаются друг с другом
Нейроны общаются друг с другом через синапсов . Когда потенциал действия достигает пресинаптического окончания, он вызывает высвобождение нейротрансмиттера из нейрона в синаптическую щель , зазор в 20–40 нм между окончанием синаптического аксона до и синаптическим дендритом после (часто шип). .
После прохождения через синаптическую щель передатчик будет присоединяться к рецепторам нейротрансмиттера на постсинаптической стороне, и в зависимости от высвобождаемого нейромедиатора (что зависит от типа нейрона, высвобождающего его), особенно положительного (например, Na + , K + , Ca + ) или отрицательные ионы (например, Cl — ) будут проходить через каналы, проходящие через мембрану.
Синапсы можно рассматривать как преобразование электрического сигнала (потенциала действия) в химический сигнал в виде высвобождения нейротрансмиттера, а затем, после связывания передатчика с постсинаптическим рецептором, снова переключая сигнал в электрическую форму , когда заряженные ионы втекают или выходят из постсинаптического нейрона.
Потенциал действия, или спайк, вызывает высвобождение нейротрансмиттеров через синаптическую щель, вызывая электрический сигнал в постсинаптическом нейроне. (Изображение: Томас Сплеттстессер / CC BY-SA 4.0)
Видео: потенциалы действия в нейронах
Понятия и определения
Аксон — длинная тонкая структура, в которой генерируются потенциалы действия; передающей части нейрона. После инициации потенциалы действия распространяются по аксонам, вызывая высвобождение нейротрансмиттера.
Дендрит – Принимающая часть нейрона. Дендриты получают синаптические входы от аксонов, при этом общая сумма дендритных входов определяет, будет ли нейрон запускать потенциал действия.
Позвоночник — Небольшие выступы на дендритах, которые для многих синапсов являются местом постсинаптического контакта.
Мембранный потенциал — Электрический потенциал через клеточную мембрану нейрона, возникающий из-за различного распределения положительно и отрицательно заряженных ионов внутри и снаружи клетки. Значение внутри ячейки всегда указывается относительно внешнего: -70 мВ означает, что внутреннее значение на 70 мВ более отрицательное, чем внешнее (которому присваивается значение 0 мВ).
Потенциал действия — Кратковременное (~ 1 мс) электрическое событие, обычно генерируемое в аксоне, которое сигнализирует нейрону об «активности». Потенциал действия распространяется по длине аксона и вызывает высвобождение нейротрансмиттера в синапс. Потенциал действия и последующее высвобождение медиатора позволяют нейрону связываться с другими нейронами.
Нейротрансмиттер — Химическое вещество, высвобождаемое нейроном после потенциала действия. Нейротрансмиттер проходит через синапс, возбуждая или подавляя нейрон-мишень. Различные типы нейронов используют разные нейротрансмиттеры и, следовательно, по-разному воздействуют на свои мишени.
Синапс — соединение между аксоном одного нейрона и дендритом другого, через которое два нейрона сообщаются друг с другом.
Исследования QBI
Лаборатории QBI, работающие над нейронами и нейронной коммуникацией: профессор Стивен Уильямс, профессор Панкадж Сах
Лаборатории QBI, работающие над синапсами: доктор Виктор Анггоно, профессор Джозеф Линч, профессор Фредерик Менье
Потенциал действия | Биология для специальностей II
Результаты обучения
- Объяснять этапы потенциала действия и то, как распространяются потенциалы действия
Нейрон может получать входные данные от других нейронов и, если эти входные данные достаточно сильны, посылать сигнал нижестоящим нейронам. Передача сигнала между нейронами обычно осуществляется химическим веществом, называемым нейротрансмиттером. Передача сигнала внутри нейрона (от дендрита к окончанию аксона) осуществляется за счет кратковременного изменения мембранного потенциала покоя, называемого потенциал действия . Когда молекулы нейротрансмиттера связываются с рецепторами, расположенными на дендритах нейрона, открываются ионные каналы. В возбуждающих синапсах это открытие позволяет положительным ионам проникать в нейрон и приводит к деполяризации мембраны — уменьшению разницы в напряжении между внутренней и внешней частью нейрона. Стимул от сенсорной клетки или другого нейрона деполяризует нейрон-мишень до его порогового потенциала (-55 мВ). Каналы Na + в аксонном холмике открываются, позволяя положительным ионам проникать в клетку (рис. 1).
Как только открываются натриевые каналы, нейрон полностью деполяризуется до мембранного потенциала около +40 мВ. Потенциалы действия считаются событием «все или ничего», поскольку при достижении порогового потенциала нейрон всегда полностью деполяризуется. Как только деполяризация завершена, клетка должна теперь «сбросить» свой мембранный потенциал обратно к потенциалу покоя. Для этого каналы Na + закрываются и не могут быть открыты. Так начинается рефрактерный период нейрона 90 930 9.0931, в котором он не может произвести другой потенциал действия, потому что его натриевые каналы не открываются. В то же время потенциалзависимые каналы K + открываются, позволяя K + покинуть клетку. Когда ионы К + покидают клетку, мембранный потенциал снова становится отрицательным. Диффузия K + из клетки на самом деле гиперполяризует клетку в том смысле, что мембранный потенциал становится более отрицательным, чем нормальный потенциал покоя клетки. В этот момент натриевые каналы вернутся в состояние покоя, то есть готовы снова открыться, если мембранный потенциал снова превысит пороговый потенциал. В конце концов дополнительный K 9Ионы 0172 + диффундируют из клетки через каналы утечки калия, возвращая клетку из ее гиперполяризованного состояния обратно к мембранному потенциалу покоя.
Практический вопрос
Формирование потенциала действия можно разделить на пять этапов, которые показаны на рисунке 1.
Рисунок 1. Потенциал действия
- Стимул от сенсорной клетки или другого нейрона вызывает целевую клетку деполяризоваться в сторону порогового потенциала.
- При достижении порога возбуждения открываются все каналы Na + и мембрана деполяризуется.
- На пике потенциала действия открываются каналы К + , и К + начинают покидать клетку. При этом закрываются каналы Na + .
- Мембрана становится гиперполяризованной по мере того, как ионы K + продолжают покидать клетку. Гиперполяризованная мембрана находится в рефрактерном периоде и не может сработать.
- Каналы K + закрываются и Na + /K + транспортер восстанавливает потенциал покоя.
Блокаторы калиевых каналов, такие как амиодарон и новокаинамид, которые используются для лечения аномальной электрической активности в сердце, называемой сердечной аритмией, препятствуют движению K + через потенциалзависимые каналы K + . На какую часть потенциала действия, как вы ожидаете, повлияют калиевые каналы?
Показать ответ
Рисунок 2. Потенциал действия проводится вниз по аксону по мере того, как мембрана аксона деполяризуется, а затем реполяризуется.
В этом видео представлен обзор потенциала действия.
Миелин и распространение потенциала действия
Чтобы потенциал действия передал информацию другому нейрону, он должен пройти вдоль аксона и достичь окончаний аксона, где он может инициировать высвобождение нейротрансмиттера. На скорость проведения потенциала действия по аксону влияет как диаметр аксона, так и сопротивление аксона утечке тока. Миелин действует как изолятор, который предотвращает выход тока из аксона; это увеличивает скорость проведения потенциала действия. При демиелинизирующих заболеваниях, таких как рассеянный склероз, проведение потенциала действия замедляется из-за утечки тока из ранее изолированных областей аксонов.
Рисунок 3. Узлы Ранвье представляют собой разрывы в покрытии миелина вдоль аксонов. Узлы содержат потенциалзависимые каналы K + и Na + . Потенциалы действия распространяются по аксону, перескакивая с одного узла на другой.
Перехваты Ранвье, показанные на рисунке 3, представляют собой разрывы в миелиновой оболочке вдоль аксона. Эти немиелинизированные пространства имеют длину около одного микрометра и содержат потенциалзависимые каналы Na + и K + . Поток ионов через эти каналы, особенно Na + каналов, снова и снова регенерирует потенциал действия вдоль аксона. Этот «перескок» потенциала действия от одного узла к другому называется скачкообразной проводимостью . Если бы перехваты Ранвье не присутствовали вдоль аксона, потенциал действия распространялся бы очень медленно, поскольку каналы Na + и K + должны были бы непрерывно регенерировать потенциалы действия в каждой точке вдоль аксона, а не в определенных точках. Узлы Ранвье также экономят энергию нейрона, поскольку каналы должны присутствовать только в узлах, а не вдоль всего аксона.
Попробуйте
Внесите свой вклад!
У вас есть идеи по улучшению этого контента? Мы будем признательны за ваш вклад.
Улучшить этот Pagelearn More
Потенциал действия — Потенциал мембраны покоя — Потенциал генерации действий
- 1 Потенциал мембраны покоя
- 2 Генерация Действия
- 3 Процесс потенциалов действия
- 3,1 MyELELATED AXSON
- 4 Клиническая значимость — рассеянный склероз
Нейроны общаются друг с другом с помощью коротких электрических сигналов, известных как потенциалы действия . Это кратковременные изменения напряжения на мембране из-за потока определенных ионов в нейрон и из него. В этой статье мы обсудим, как генерируется потенциал действия (ПД) и как происходит проведение потенциала действия.
Потенциал мембраны покоя
Потенциал мембраны покоя клеток различается в зависимости от типа клеток.
Для нейронов он обычно находится между -50 и -75 мВ. Это значение зависит от типа открытых ионных каналов и концентрации различных ионов во внутриклеточной и внеклеточной жидкости в состоянии покоя. В нейронах K+ и органические анионы обычно находятся в более высокой концентрации внутри клетки, чем снаружи, тогда как Na+ и Cl- обычно находятся в более высоких концентрациях вне клетки.Эта разница в концентрациях обеспечивает градиент концентрации для стекания ионов вниз, когда их соответствующие каналы открыты. Следовательно, ионы K+ будут перемещаться из клеток, а ионы Na+ и Cl- будут перемещаться в клетку. В состоянии покоя клетка в основном проницаема для K+, поэтому он оказывает наибольшее влияние на мембранный потенциал покоя из трех ионов. Дополнительную информацию о генерации потенциала покоя можно найти здесь.
Эти градиенты концентрации поддерживаются действием Na+/K+ АТФазы посредством активного транспорта, что, в свою очередь, позволяет поддерживать мембранный потенциал.
Synaptidude [CC BY 3.0 (https://creativecommons.org/licenses/by/3.0)], с Wikimedia Commons
градиенты концентрации ионов.
Генерация потенциалов действия
В состоянии покоя возникает мембранный потенциал, так как мембрана преимущественно проницаема для K+. Потенциал действия начинается на аксонном холмике в результате деполяризации. Во время деполяризации потенциалзависимые ионные каналы натрия открываются из-за электрического стимула. Когда ионы натрия устремляются обратно в клетку, их положительный заряд меняет потенциал внутри клетки с отрицательного на более положительный.
Если достигается пороговый потенциал , возникает потенциал действия. Потенциалы действия возникают только при достижении порога. Кроме того, если порог достигнут, всегда вызывается ответ одинаковой величины, независимо от силы раздражителя. Следовательно, потенциалы действия описываются как « все или ничего ».
Как только клетка деполяризуется, потенциал-управляемые натриевые ионные каналы начинают закрываться. Положительный потенциал внутри клетки вызывает открытие потенциалзависимых калиевых каналов , и теперь ионы K+ перемещаются по своему электрохимическому градиенту из клетки. Когда K+ выходит из клетки, мембранный потенциал становится более отрицательным и начинает приближаться к потенциалу покоя.
Обычно реполяризация превышает мембранный потенциал покоя, делая мембранный потенциал более отрицательным. Это известно как гиперполяризация. Важно отметить, что Na+/K+ АТФаза не участвует в процессе реполяризации после потенциала действия.
За каждым потенциалом действия следует рефрактерный период. Этот период можно разделить на:
- период абсолютной рефрактерности , который наступает после закрытия натриевых каналов после ПД. Затем натриевые каналы переходят в неактивное состояние , во время которого они не могут быть вновь открыты, независимо от мембранного потенциала.
и
- относительный рефрактерный период , который возникает, когда натриевые каналы медленно выходят из состояния инактивации. В этот период нейрон может возбуждаться стимулами более сильными, чем те, которые обычно необходимы для инициации ПД. В начале периода относительной рефрактерности сила требуемого стимула очень высока. Постепенно он становится меньше в течение относительного рефрактерного периода по мере того, как больше натриевых каналов восстанавливается после стадии инактивации.
Автор оригинала en:User:Chris 73, обновлено en:User:Diberri, преобразовано в SVG автором tiZom [GFDL (http://www.gnu.org/copyleft/fdl.html)
Рис. 2 – Диаграмма показывая фазы потенциала действия по отношению к мембранному напряжению во времени.
Распространение потенциалов действия
Потенциалы действия распространяются по аксонам нейронов посредством локальных токов . Локальные токи вызывают деполяризацию соседней аксональной мембраны, и там, где она достигает порога, генерируются дополнительные потенциалы действия.
Участки мембраны, которые недавно деполяризовались, больше не деполяризуются из-за рефрактерного периода, т. е. 9Потенциал действия 0037 будет распространяться только в одном направлении. Эти локальные токи в конечном итоге будут уменьшать заряд до тех пор, пока пороговое значение больше не будет достигнуто. Расстояние, которое это займет, зависит от емкости и сопротивления мембраны:
- Емкость мембраны – способность накапливать заряд. Более низкая емкость приводит к большему расстоянию до того, как пороговое значение больше не будет достигнуто.
- Сопротивление мембраны – зависит от количества открытых ионных каналов. Чем меньше количество открытых каналов, тем больше сопротивление мембраны. Более высокое сопротивление мембраны приводит к большему расстоянию до того, как порог больше не будет достигнут.
Джон Шмидт (Марс) [GFDL (http://www.gnu.org/copyleft/fdl.html)
Рис. 3 — Анимация, показывающая, как потенциал действия распространяется по аксону.
Миелиновые аксоны
Чтобы обеспечить быструю передачу электрических сигналов через нейрон и сделать их более энергоэффективными, некоторые аксоны нейронов покрыты миелиновой оболочкой . Миелиновая оболочка окружает аксон, образуя изолирующий слой. Дополнительную информацию о миелиновой оболочке можно найти здесь.
Вдоль миелинизированного аксона имеются периодические щели, в которых отсутствует миелин и обнажается аксональная мембрана. Эти пробелы называются N оды Ранвье. В отличие от миелинизированных участков аксона, в которых отсутствуют потенциалзависимые ионные каналы, перехваты Ранвье содержат высокую плотность ионных каналов. По этой причине потенциал действия может возникать только в узлах.
Миелиновая оболочка ускоряет проводимость за счет увеличения сопротивления мембраны и уменьшение емкости мембраны. Следовательно, потенциал действия может распространяться по нейрону с более высокой скоростью, чем это было бы возможно в немиелинизированных нейронах.
Электрические сигналы быстро передаются от одного узла к другому, что вызывает деполяризацию мембраны. Если деполяризация превышает порог, она инициирует другой потенциал действия, который проводится к следующему узлу. Таким образом, потенциал действия быстро передается по нейрону. Это известно как скачкообразная проводимость. Автор Helixitta [CC BY-SA 4.0 (https://creativecommons.org/licenses/by-sa/4.0)], с Wikimedia Commons
Рис. 4. Диаграмма, показывающая, как миелиновая оболочка приводит к скачкообразной проводимости потенциал действия вдоль аксона.
Клиническая значимость — рассеянный склероз
Рассеянный склероз (РС) — приобретенное хроническое аутоиммунное заболевание, поражающее ЦНС. Это приводит к демиелинизации, глиозу и повреждению нейронов. Обычными проявлениями болезни являются неврит зрительного нерва, поперечный миелит и мозжечковые симптомы, такие как атаксия.
Микаэль Хэггстрем, используется с разрешения.
(Все использованные изображения находятся в общественном достоянии.) [Общественное достояние], через Wikimedia CommonsРис. 5. Диаграмма, демонстрирующая основные симптомы рассеянного склероза.
Различают три основные формы заболевания:
- Ремиттирующе-рецидивирующее – Пациенты сталкиваются с эпизодами ремиссии (во время которых симптомы отсутствуют) и обострениями заболевания.
- Вторичный прогрессивный — Первоначально рассеянный склероз имеет рецидивирующе-ремиттирующий характер. Однако в какой-то момент течение заболевания меняется, и неврологическая функция постепенно ухудшается.
- Первично-прогрессирующий – После начала заболевания наблюдается неуклонное прогрессирование и ухудшение течения болезни.
Лекарство от рассеянного склероза не известно. Однако некоторые методы лечения оказались полезными с точки зрения лечения острых обострений, предотвращения обострений и замедления инвалидности.
Например, высокие дозы внутривенного кортикостероиды могут помочь облегчить симптомы при острых обострениях.print Print this Article
Потенциалы действия имеют решающее значение для распространения локально вызванных распространяющихся деполяризаций
Сохранить цитату в файл
Формат: Резюме (текст)PubMedPMIDAbstract (текст)CSV
Добавить в коллекции
- Создать новую коллекцию
- Добавить в существующую коллекцию
Назовите свою коллекцию:
Имя должно содержать менее 100 символов
Выберите коллекцию:
Не удалось загрузить вашу коллекцию из-за ошибки
Повторите попыткуДобавить в мою библиографию
- Моя библиография
Не удалось загрузить делегатов из-за ошибки
Повторите попыткуВаш сохраненный поиск
Название сохраненного поиска:
Условия поиска:
Тестовые условия поиска
Эл.
адрес:
(изменить)Который день? Первое воскресеньеПервый понедельникПервый вторникПервая средаПервый четвергПервая пятницаПервая субботаПервый деньПервый будний день
Который день? ВоскресеньеПонедельникВторникСредаЧетвергПятницаСуббота
Формат отчета: SummarySummary (text)AbstractAbstract (text)PubMed
Отправить максимум: 1 шт. 5 шт. 10 шт. 20 шт. 50 шт. 100 шт. 200 шт.
Отправить, даже если нет новых результатов
Необязательный текст в электронном письме:
Создайте файл для внешнего программного обеспечения для управления цитированием
Полнотекстовые ссылки
HighWire
Полнотекстовые ссылки
.
2022 16 марта; 42 (11): 2371-2383.doi: 10.1523/JNEUROSCI.2930-20.2021. Epub 2021 2 декабря.
Пратюш Сурьяванши 1 2 , Кэтлин М Рейнхарт 1 , К. Уильям Шаттлворт 3 , К. С. Бреннан 4
Принадлежности
- 1 Кафедра неврологии, Медицинский факультет Университета Юты, Солт-Лейк-Сити, Юта 84108.
- 2 Межведомственная программа нейробиологии, Медицинский факультет Университета Юты, Солт-Лейк-Сити, Юта 84108.
- 3 Отделение неврологии, Медицинский факультет Университета Нью-Мексико, Альбукерке, Нью-Мексико 87131.
- 4 Кафедра неврологии, Медицинский факультет Университета Юты, Солт-Лейк-Сити, Юта 84108 [email protected].
- PMID: 34857650
- PMCID: PMC8936615
- DOI: 10.1523/JNEUROSCI.2930-20.2021
Бесплатная статья ЧВК
Пратюш Сурьяванши и др. Дж. Нейроски. .
Бесплатная статья ЧВК
. 2022 16 марта; 42 (11): 2371-2383.
doi: 10.1523/JNEUROSCI.2930-20.2021. Epub 2021 2 декабря.
Авторы
Пратюш Сурьяванши 1 2 , Кейтлин М Рейнхарт 1 , К. Уильям Шаттлворт 3 , К. С. Бреннан 4
Принадлежности
- 1 Кафедра неврологии, Медицинский факультет Университета Юты, Солт-Лейк-Сити, Юта 84108.
- 2 Межведомственная программа нейробиологии, Медицинский факультет Университета Юты, Солт-Лейк-Сити, Юта 84108.
- 3 Отделение неврологии, Медицинский факультет Университета Нью-Мексико, Альбукерке, Нью-Мексико 87131.
- 4 Кафедра неврологии, Медицинский факультет Университета Юты, Солт-Лейк-Сити, Юта 84108 [email protected].
- PMID: 34857650
- PMCID: PMC8936615
- DOI: 10.1523/JNEUROSCI.2930-20.2021
Абстрактный
Распространяющиеся деполяризации (СД) серого вещества возникают в головном мозге при различных патологических состояниях и вызывают разную степень повреждения тканей в зависимости от степени метаболической нагрузки на ткани. Как и следовало ожидать для таких больших деполяризаций, нейроны демонстрируют всплески потенциалов действия (ПД) по мере распространения волны.
Однако конкретная роль AP в распространении SD неясна. Это потенциально важно, поскольку модуляция натриевых каналов не рассматривалась в качестве терапевтической мишени для расстройств, связанных с SD, из-за неоднозначных экспериментальных данных. Используя электрофизиологию целых клеток и однофотонную визуализацию острых срезов коры головного мозга самцов мышей C57Bl6, мы проверили влияние блокады AP на SD, генерируемые двумя широко используемыми индукционными парадигмами. Мы обнаружили, что блокада ПД с использованием тетродотоксина (ТТХ) ограничивала распространение фокально индуцированных СД и значительно уменьшала амплитуду деполяризации нейронов, а также ее Са 2+ нагрузка. TTX также устранял подавление спонтанной синаптической активности, которая является отличительной чертой фокально индуцированной SD. Напротив, TTX не влиял на распространение SD, вызванное глобальной суперфузией высоко [K + ] e , содержащих искусственный CSF (ACSF). Таким образом, мы показываем, что опосредованные потенциал-зависимыми натриевыми каналами (Na v ) всплески нейронных AP имеют решающее значение для распространения и нижестоящих эффектов фокально индуцированной SD, но менее важны, когда ионный баланс внеклеточного пространства уже нарушен. При этом мы подтверждаем мнение о том, что две разные парадигмы индукции СЗ, каждая из которых относится к разным клиническим ситуациям, значительно различаются по своим характеристикам и, возможно, по реакции на лечение. ЗАЯВЛЕНИЕ О ЗНАЧИМОСТИ Наши результаты показывают, что потенциалзависимые натриевые каналы (Na v ) играют критическую роль в распространении и нижестоящих нервных эффектах фокально индуцированной распространяющейся деполяризации (SD). Поскольку СЗ, вероятно, индуцируются локально при многих болезненных состояниях, эти исследования поддерживают модуляцию натриевых каналов, ранее недооцениваемый терапевтический вариант при расстройствах, связанных с СЗ, как жизнеспособный подход.Ключевые слова: Травма головного мозга; мигрень; потенциалы действия нейронов; возбудимость нейронов; распространение деполяризации; распространяющаяся депрессия.
Copyright © 2022 авторы.
Цифры
Рисунок 1.
Индукция SD с использованием фокусного и…
Рисунок 1.
Индукция SD с использованием фокальных и глобальных стимулов зажигания. A , слева, Пример изображения…
Фигура 1.Индукция SD с использованием фокальных и глобальных стимулов зажигания. A , слева, пример изображения, показывающий поток ACSF и размещение пипетки с затяжкой (выталкиванием под давлением). Справа репрезентативные изображения (оттенки серого), показывающие острое распространение SR101 (излучение: >605 нм) после однократной затяжки (0,5 бар N 2 давление, устойчивость к пипетке 05–0,8 МОм, шкала: 200 мкм). B , Количественная оценка площади сигнала SR101, через 5 мин после однократной затяжки.
C , Пример последовательности изображений (ΔT), показывающий развитие одиночной очаговой SD, вызванной 3-метровым выбросом K + под давлением (вверху), а также распространение двух волн SD, вызванных глобальным 40 мм K + перфузии (внизу). D . Схема, показывающая круговую область интереса (область интереса 1, диаметр 50 мкм), а также линейную область интереса (область интереса 2, длина 1337 мкм), расположенную на кортикальном уровне L2/3, для захвата SD, генерируемого фокальным высоким [К + ] слойка. E , вверху, пример кривой (ΔT/T 0 ), созданной с использованием круговой области интереса, показывающей изменение IOS во время SD в определенном месте коры L/23. Внизу: кимограф, показывающий временную прогрессию изменения IOS, связанного с SD, во времени, созданный с использованием линейных ROI (расстояние в зависимости от времени). Скорость распространения SD количественно определяли как наклон изменения IOS на кимографе через фиксированные интервалы, представленные тремя равноудаленными отрезками линии (400 мкм). F , Количественное определение уклонов ((y 2 – y 1 )/(x 2 – x 1 )) в виде скоростей распространения (мм/мин) для соответствующих сегментов расстояния. G . Схема, показывающая создание линейной области интереса по L2/3 для захвата однократных отклонений, генерируемых высокой суперфузией [K + ]. H , Кимограмма (расстояние в зависимости от времени), полученная из области интереса, показывающая временное развитие двух однонаправленных волн, распространяющихся с противоположных направлений.Рисунок 2.
Увеличение [K + ]…
Рисунок 2.
Увеличение [K + ] e , совпадающее с распространением фокально индуцированного SD.
Фигура 2. …Увеличение [K + ] e , совпадающее с распространением фокально индуцированной SD. A . Схема, показывающая фокальную индукцию SD с использованием выброса давления на 3 м [K + ] и одновременное получение IOS (визуализация) вместе с [K + ] e и запись постоянного тока с использованием K + — чувствительный биполярный микроэлектрод (см. Материалы и методы). B , Калибровочная кривая, показывающая изменение напряжения (мВ), зарегистрированное с помощью селективного электрода K + в ответ на увеличение концентрации K + ( R 2 = 0,9733, наклон = 50,14, n = 3 электрода). C , Пик [K + ] e в месте регистрации при распространении СД не коррелирует с расстоянием от индукции ( p = 0,58, корреляция Пирсона r = -91,23, n = 8).
D , вверху, последовательность изображений, показывающая размещение индукционной пипетки K + (обозначена *), K + -чувствительный микроэлектрод (обозначен стрелкой), а также размещение ROI вместе с распространением SD в реальном времени (увеличение ΔT) в течение 10 с после индукции. Электрод и ROI располагались на расстоянии ~474 мкм от места индукции. После индукции СД распространялась в направлении K + -электрод чувствительный. Внизу репрезентативная кривая, показывающая реакцию напряжения на увеличение внеклеточного K + (K + — чувствительный электрод), а также сигнал постоянного тока (электрод сравнения). [K + ] e в этом месте не повышалось во время индукции SD (фокальное K + затяжка), но увеличивалось одновременно с сигналом DC и ROI с пиками всех трех сигналов, выровненными во времени. E , вверху, последовательность изображений, показывающая расположение K + индукционная пипетка (затяжка), K + — чувствительный микроэлектрод, а также размещение ROI, 376 мкм от места индукции. Увеличение ΔT после индукции SD показывает распространение SD от электрода, чувствительного к K + . Внизу репрезентативная кривая, показывающая отсутствие изменений во внеклеточном сигнале K + , DC или IOS ни во время, ни после индукции SD, что подтверждает наши наблюдения роста [K + ] e только во время распространения SD.Рисунок 3.
Определенная кинетика и продолжительность подъема…
Рисунок 3.
Различная кинетика нарастания и продолжительность деполяризации нейронов, вырожденных двумя разными…
Рисунок 3.Различная кинетика нарастания и продолжительность деполяризации нейронов, вырожденных двумя различными парадигмами индукции SD.
A . Схема, показывающая индукцию SD с использованием выброса фокального давления на 3 м [K + ] (слева), вместе с репрезентативным следом нейрона V м во время SD-волны (зажим I ), зарегистрированным > 500 мкм от места индукции. B . Схема, показывающая индукцию SD с использованием глобальной суперфузии 40 мм [K + ] (слева), наряду с репрезентативным следом нейронов V m во время SD. Нет разницы в амплитуде пика ( C , p = 0,14, критерий Манна-Уитни), а также абсолютный пик V m ( D , p = 0,98, критерий Манна-Уитни) индукционные парадигмы. E , Общая продолжительность деполяризации SD была достоверно выше при индукции 40 мм К + ACSF ( p = 0,016, критерий Манна-Уитни), без достоверной разницы в продолжительности вспышек ПД ( Ф , p = 0,86, критерий Манна–Уитни) между индукционными парадигмами. G , Линия линейной регрессии, подогнанная к нейронным трассам V m , показывающая разные наклоны подъема SD для двух различных парадигм индукции. Время до пика было значительно уменьшено (вставка, p = 0,0002, критерий Манна-Уитни) для SD, вызванных фокальной 3-метровой затяжкой K + . (* P < 0,05, *** P < 0,001).Рисунок 4.
Отчетливая кинетика подъема IOS…
Рисунок 4.
Отчетливая кинетика нарастания сигнала IOS после индукции SD двумя парадигмами.…
Рисунок 4.Отчетливая кинетика нарастания сигнала IOS после индукции SD двумя парадигмами. A , последовательность изображений IOS (ΔT), показывающая прогрессирование SD, индуцированного 3 м K + выброс под давлением (вверху), а также перфузия 40 мм K + ACSF (внизу) в пространстве и времени (масштабная линейка: 200 мкм).
B , Кривые, показывающие изменения коэффициента пропускания (ΔT/T 0 , групповые средние выделены жирным шрифтом) во время прохождения однонаправленной волны (ROI ~1000 мкм от места индукции). C , Линия линейной регрессии, подогнанная к средним значениям ΔT/T 0 , показывает различные наклоны подъема SD для двух различных парадигм индукции. D , Максимальное зарегистрированное распространение однонаправленных волн, вызванных двумя различными парадигмами индукции, не отличается (непарный тест t , p = 0,53). E , Хотя сравнение скоростей распространения между двумя парадигмами индукции не было значимым ( p = 0,98, двусторонний дисперсионный анализ), скорость распространения значительно уменьшалась с расстоянием для фокально индуцированного SD ( p < 0,01, критерий Краскела-Уоллиса).Рисунок 5.
Обработка ТТХ ограничивает распространение…
Рисунок 5.
Обработка TTX ограничивает распространение SD, вызванного фокальным высоким [K + ]…
Рисунок 5. ЛечениеTTX ограничивает распространение SD, вызванного фокальным высоким [K + ] слойка. A . Схема, показывающая индукцию SD с использованием фокального выброса давления на 3 м [K + ] (слева) вместе с репрезентативными трассами сигнала пропускания (ΔT/T 0 ), показывающими распространение SD в условиях контроля и TTX ( ROI: 400, 800 и 1200 мкм от места индукции). B , Пример Последовательность изображений (ΔT), показывающая распространение фокально индуцированной SD-волны в пространстве и времени в условиях контроля, а также в условиях лечения TTX (масштабная линейка: 200 мкм).
C , Кривые, показывающие изменения коэффициента пропускания света (ΔT/T 0 ) после фокально-индуцированного SD, в контрольных ( n = 9, синий) и TTX ( n = 11, серый) условиях (ROI) при ∼500 и 1000 мкм групповые средние выделены жирным шрифтом). D , обработка ТТХ значительно уменьшила максимальное распространение SD волны, генерируемой фокальным выбросом давления на 3 м [K + ] (непарный тест t , p <0,001), не влияя на скорость SD ( E ) между двумя группами (двусторонний ANOVA, p = 0,07). (*** P < 0,001).Рисунок 6.
Распространение SD, генерируемое…
Рисунок 6.
На распространение SD, генерируемого глобальным максимумом [K + ], не влияет…
Рисунок 6. Распространение SD, генерируемого глобальным максимумом [K + ], не зависит от обработки TTX. A . Схема, показывающая индукцию SD с использованием глобальной суперфузии 40 мм [K + ] (слева) вместе с репрезентативными кривыми сигнала пропускания (ΔT/T 0 ), показывающими распространение SD в контроле и условиях TTX (ROI) : 400, 800 и 1200 мкм от места индукции). B , Пример последовательности изображений (ΔT), показывающий продвижение общего 40 мм [K + ] индуцированная суперфузией SD-волна в пространстве и времени в контроле, а также в условиях лечения TTX (масштабная линейка: 200 мкм). C , Кривые, показывающие изменения коэффициента пропускания света (ΔT/T 0 ) в ответ на SD, вызванные 40 мм [K + ] суперфузией, в контроле (вверху, n = 10, красный) как а также условия TTX (внизу, n = 11, серый) (ROI ~ 1000 мкм, средние значения группы выделены жирным шрифтом).
В случае SD, вызванного перфузией 40 мм K + ACSF, обработка TTX не влияла ни на максимальное распространение ( D , непарный тест t , p = 0,08), или SD-скорость ( E , двухсторонний ANOVA, p = 0,86).Рисунок 7.
Лечение ТТХ сокращает продолжительность…
Рисунок 7.
Терапия ТТХ уменьшает продолжительность и амплитуду ДС, отдаленных от индукции…
Рисунок 7. ЛечениеТТХ уменьшает продолжительность и амплитуду СЗ, удаленных от места индукции. A . Схема, показывающая фокальную индукцию SD с использованием выброса давления на 3 м [K + ] и сбор данных с использованием фиксации тока всей клетки (слева), а также типичные следы фиксации тока для SD в контроле (синий) и Условия ТТХ (серые) (стрелки указывают на высокий [K + ] слой).
Терапия ТТХ значительно уменьшала продолжительность ДС ( B , Mann–Whitney test, p = 0.043), maximum depolarization ( C , Mann–Whitney test, p < 0.0038) and peak amplitude ( D , Mann– Уитни, p = 0,0024), а также отменили всплески ПД при СД ( E , критерий Манна-Уитни, p < 0,0001, контроль N = 9, ТТХ N = 9). F . Схема, показывающая записи токовых зажимов фокально индуцированной SD, полученные от проксимальных (~200 мкм) и дистальных (~800 мкм) нейронов в острых срезах. G , Репрезентативные следы деполяризации V m , связанной с SD, в присутствии TTX в нейронах проксимальнее (вверху ~ 200 мкм) и дистальнее (внизу ~ 800 мкм) от места индукции. H , Отрицательная корреляция между амплитудой SD и расстоянием от места индукции (непараметрический коэффициент корреляции Спирмена r S = -0,9429, p = 0,017). I , Репрезентативные следы подъема V м (отфильтрованы для исключения AP) во время SD, демонстрирующие многофазный подъем с двумя постоянными времени в контрольных условиях и экспоненциальный рост с одной постоянной времени в условиях TTX. J , Сравнение постоянных времени (τ 1 ) однофазных экспоненциальных подгонок к начальному подъему V м (предшествующих всплескам ПД) между контролем и условиями ТТХ не выявило значимой разницы ( p = 0,95 , критерий Манна–Уитни, контроль n = 11, ТТХ n = 7). K , Сравнение времени до пика между контрольной группой и группой TTX не показало существенной разницы ( p = 0,53, критерий Манна-Уитни, контроль н =11, ТТХ н =7). (* P < 0,05, ** P < 0,01, *** P < 0,001).Рисунок 8.
Внутриклеточная кальциевая нагрузка, связанная с SD, составляет…
Рисунок 8.
Внутриклеточная кальциевая нагрузка, связанная с SD, значительно снижается при лечении TTX. А , Схема…
Рисунок 8.Внутриклеточная кальциевая нагрузка, связанная с SD, значительно снижается при лечении TTX. A . Схема, показывающая фокальную индукцию SD наряду с эпифлуоресцентной визуализацией в острых срезах, полученных от животных Thy1-GCamp3. B , Репрезентативные следы флуоресцентного сигнала (ΔF/F 0 ), показывающие распространение SD в контрольных срезах и срезах, обработанных TTX (ROI: 400, 800 и 1200 мкм от места индукции). C , Пример последовательности изображений (ΔF), показывающий распространение фокально индуцированной SD-волны (3 м K + выброс давления ) в пространстве и времени в контрольных условиях и условиях TTX (масштабная линейка: 200 мкм). D , Кривые, показывающие изменения сигнала флуоресценции (ΔF/F 0 ) в ответ на фокально индуцированный SD, зарегистрированный для контроля (вверху, n = 8, синий), а также ТТХ (внизу, n = 7, серый) срезы (ROI на расстоянии ~ 500 и 1000 мкм от места индукции).
E , TTX не влиял на скорость распространения SD-ассоциированного сигнала Ca 2+ ( p = 0,65, двусторонний ANOVA). F , Однако лечение ТТХ значительно уменьшило максимальное расстояние распространения фокально генерируемой SD волны ( p = 0,023, критерий Манна-Уитни). G , Аналогично, площадь под кривой (ΔF/F 0 *время), которая соответствует нагрузке ткани Ca 2+ , также уменьшается после обработки ТТХ как при 500 мкм ( p = 0,031, непарный t тест), а также 1000 мкм ( p = 0,024, непарный t тест). (* P < 0,05, ** P < 0,01).Рисунок 9.
Обработка TTX предотвращает индуцированную SD…
Рисунок 9.
Лечение TTX предотвращает индуцированное SD подавление возбуждающей спонтанной синаптической передачи.
Рисунок 9. Лечение А ,…TTX предотвращает индуцированное SD подавление возбуждающей спонтанной синаптической передачи. A , репрезентативные кривые, показывающие мВПСК, зарегистрированные до (базовый уровень), через 5 минут после и через 30 минут после фокально индуцированного ДД в присутствии ТТХ. B , Сравнение средних частот мВПСТ отдельных нейронов не показывает изменений по сравнению с исходным уровнем ( стр. = 0,87, парный критерий Уилкоксона). C , Гистограмма кумулятивной частоты интервалов между событиями не показывает разницы в частоте мВПСТ между исходным уровнем и 5 мин ( p = 0,061, двухвыборочный тест KS) и 30 мин ( p = 0,45, двухвыборочный KS тест) после SD ( n = 4 нейрона для амплитуды и частоты). D , Отсутствие изменений средних амплитуд мВПСТ через 5 и 30 мин после индукции SD ( p = 0,38, критерий подобранных пар Уилкоксона).
E , Гистограмма кумулятивной частоты амплитуд отдельных событий мВПСТ не выявила различий через 5 минут ( p = 0,076, двухвыборочный тест KS) и 30 минут ( p = 0,091, двухвыборочный тест KS) от базовый уровень.См. это изображение и информацию об авторских правах в PMC
Похожие статьи
Уменьшение доступности потенциал-зависимых натриевых каналов в результате деполяризации или блокады тетродотоксином усиливает взрывную активацию и высвобождение катехоламинов в хромаффинных клетках мыши.
Вандаль Д.Х., Оттавиани М.М., Легрос С., Лефор С., Герино Н.К., Аллио А., Карабелли В., Карбоне Э. Вандаль Д.Х. и др. Дж. Физиол. 2015 15 февраля; 593 (4): 905-27. doi: 10.1113/jphysiol.2014.283374. Epub 2015 26 января. Дж. Физиол. 2015. PMID: 25620605 Бесплатная статья ЧВК.
Различная роль подтипов TTX-чувствительных потенциалзависимых натриевых каналов (Na V 1) в инициации и проведении потенциала действия в ноцицепторах блуждающих дыхательных путей.
Колларик М., Сун Х., Хербстсомер Р.А., Ру Ф., Кочмалова М., Микер С.Н., Ундем Б.Дж. Колларик М. и соавт. Дж. Физиол. 2018 15 апреля; 596 (8): 1419-1432. дои: 10.1113/JP275698. Дж. Физиол. 2018. PMID: 29435993 Бесплатная статья ЧВК.
Чувствительные к тетродотоксину потенциалзависимые натриевые каналы регулируют афферентные реакции мочевого пузыря на растяжение.
Гранди Л., Эриксон А., Колдуэлл А., Гарсия-Карабальо С., Рычков Г., Харрингтон А., Брайерли С.М. Гранди Л. и др. Боль. 2018 дек;159(12): 2573-2584. doi: 10.1097/j.pain.0000000000001368. Боль. 2018. PMID: 30157135
Сравнительный анализ распространения деполяризации в срезах головного мозга, подвергнутых осмотическому или метаболическому стрессу.